|
|
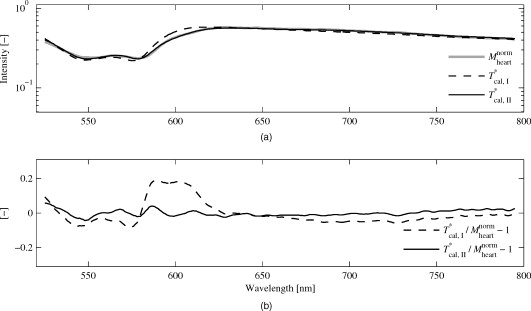
1.IntroductionA common reason for myocardial ischemia in heart patients is atherosclerotic coronary heart disease. Treatment often involves medication and/or surgical interventions, such as coronary stent implants and coronary artery bypass grafting (CABG). During the CABG procedure, the blood flow in the grafted vessels can be monitored during the surgery with ultrasound techniques.1, 2, 3 However, for proper contractility in the myocardial cells, not only adequate coronary blood flow and release of hemoglobin-bound oxygen in capillaries, but also sufficient mitochondrial oxygen uptake has to be ensured.4, 5, 6 Optical steady-state diffuse reflectance spectroscopy (DRS) has been used in several in vitro studies to study myocardial tissue status via spectral changes that are linked to chromophores involved in myocardial oxygen transport.7, 8, 9, 10 A chromophore that reveals information about the mitochondrial oxygen uptake is the cytochrome aa3 (cyt aa3), also known as cytochrome c oxidase. This is an enzyme found in the mitochondrial inner wall and is the final electron transporter in the mitochondrial respiratory chain.11 A reduction of the cyt aa3 oxidation status causes a decrease in cellular adenosine triphosphate (ATP) production. Therefore, monitoring of cyt aa3 oxidation status can be of vital importance in clinical cardiovascular medicine, since ATP production is a prerequisite for proper mechanical function in the myocardium.12 Since the optical absorbances of reduced and oxidized cyt aa3 differ, DRS is suitable for detecting changes in cyt aa3 oxidation status. However, it is a nontrivial challenge to separate the absorption effects of cyt aa3 tissue fraction from that of hemoglobin,13 although attempts have been made to measure the cyt aa3 oxidation status in the presence of hemoglobin.14 The redox state of nicotinamide adenine dinucleotide (NADH) has been described by Mayevsky and Chance as another marker in measuring mitochondrial function.15 In that study, it was concluded that the best way of monitoring the NADH redox state is by fluorometry, and that when measurements are performed on biological systems, monitoring of the NADH redox state alone is not sufficient when determining the tissue vitality. Until today, DRS techniques have relied on physiological provocations to establish reference situations involving complete oxidation and reduction of the cyt aa3,16, 17, 18 while only relative changes in the oxidation status have been measured. Obviously, extreme physiological provocations for reference measurements are not possible in clinical patient monitoring. However, publication19 of the specific extinction coefficients of cyt aa3 enables quantification of cyt aa3 tissue fraction and oxidation status without the need for physiological provocations. The aim of this study was to determine the tissue fraction and oxygenation of the myocardial hemo- and myoglobin, the tissue fraction and oxidation of cytochrome aa3, and the tissue fraction of methemoglobin during hemodynamic and respiratory provocations in calves undergoing open-chest surgery. A small intramuscular fiber-optic probe and calibrated DRS was used for measurements during variations in the oxygen fraction of inhaled gas, when unloading the heart using a left ventricular assisting device (LVAD) and, in two calves, during cardiac arrest at the end of the experiment. 2.Material and MethodsSeven calves, old, body weight ranging from , were used in this study. The animals were primarily enrolled in two different studies performed at the Animal Research Laboratory, Linköping University Hospital. Both studies were approved by the regional animal ethic committee. The main purpose of the first study (group A) was to provide open-chest situations for ablation surgery training. The main purpose of the second study (group B) was to implant and evaluate the performance of a left ventricle assisting device (LVAD) to be used in humans suffering from severe left ventricle heart failure. In both studies, the calves were premedicated with xylazine hydrochloride and atropine sulfate . A central venous catheter was inserted into the external jugular vein for administration of fluids and medication. Pentobarbital sodium was used for induction of anesthesia, and a tracheotomy was carried out to allow mechanical ventilation. Anesthesia was maintained with intravenous fentanyl and pentobarbital sodium . 2.1.Surgical TechniqueExposure of the heart was performed with a midline sternotomy. In group A, immediately after the sternotomy, the intramuscular fiber-optic probe was placed at a depth of approximately in an area of the left ventricle wall supplied by the left anterior descending (LAD) artery. The probe was fixated by epicardial suturing in order to minimize movement artifacts. In group B, extracorporeal circulation (ECC) was established after the sternotomy, and the aorta was cross-clamped together with antegrade cardioplegia infusion according to routine clinical procedures. The LVAD was implanted in the apex of the left ventricle. The aorta cross-clamp was then removed, and the LVAD was set to . The intramuscular fiber-optic probe was then positioned identically as in animals in group A. For details about the implantation, see Hübbert 20 A schematic drawing of the probe, including the dimensions, and the probe placement in both group A and group B animals are shown in Fig. 1 . Fig. 1(a) Probe design, indicating the dimensions of the probe tip. The hatched fiber indicates the emitting fiber. (b) Fiber-optic probe placement in animals from group A. (c) Fiber-optic probe placement in animals from group B, including the LVAD. The probe placement is highlighted with a white arrow.  2.2.EquipmentA multichannel spectrometer (AvaSpec 2048-5-RM, Avantes BV, Eerbeek, The Netherlands ) operating in the visible wavelength region ( , grating: VB ), was used. Access to the tissue was gained through a custom-built fiber-optic probe containing one emitting and one receiving step-index multimode fiber with a core/cladding diameter of and a numerical aperture of 0.37. The optical fibers were inserted into a slightly bent syringe cannula with a diameter of (Fig. 1, left panel). The receiving fiber was positioned adjacent to the illuminating fiber at a center–center separation of . An additional fiber, guiding light directly from the lamp to the spectrometer, was used for registering the spectrum of the light source. In-house designed algorithms and software were developed for data recording and analysis, using LabView 6.1 (National Instruments, Austin, Texas), and MATLAB 7.5 (Mathworks, Inc., Natick, Massachusetts), respectively. 2.3.Protocols and Spectral RecordingsFor each measured spectrum, a simultaneous registration of the lamp spectrum was performed. The integration time was not changed during each case, but varied from between calves. After finishing the measurements on each subject, before disconnecting the probe from the spectroscope, three reference measurements were performed. As reference, a tightly packed sample of barium sulfate powder with a reflectivity of above 98% in the visible wavelength range was used. The probe was vertically lowered approximately into the powder, allowing no surrounding light to enter. Three consecutive measurements were performed. Last, the lamp was turned off, and a dark spectrum was registered. 2.3.1.Group A (provocations with changes in )Five calves, denoted A1 to A5, were subjected to this protocol. After placement of the fiber-optic probe, at least was allowed to achieve stable heart rhythm as assessed by visual inspection of the ECG signal. In calves A1 and A2, the surgical training protocol was carried out before the spectroscopic measurements in order to enable measurements during cardiac arrest in the end of the experiment. In calves A3 to A5, measurements were performed immediately after the stabilization phase and before the surgical training. The fraction of inspired was varied in three different steps, 21%, 50%, and 100%, respectively, with duration per phase. Spectra were measured continuously during the variations. In cases A1 and A2, registrations of spectra during after initiation of cardiac arrest in the end of the experiment were also performed. 2.3.2.Group B (provocations with changes in LVAD speed and )Two calves, denoted B1 and B2, were subjected to this protocol. Similar to the group A protocol, at least of stable heart rhythm was allowed after placement of the fiber-optic probe. In calve B1, the was first held constant at 100%, while the speed of the LVAD was varied in three different steps, , , and , with duration per phase. After switching from , and from , was allowed before spectra were measured. After the duration with the LVAD speed at , the was then varied in four steps; 50%, 21%, 50%, and 100%, with duration per phase. During the different levels, spectra were measured continuously. In calf B2, the LVAD speed was held constant at throughout the experiment, and was varied in three different steps, 21%, 50%, and 100%, with duration per phase. 2.4.Spectral Preprocessing and System CalibrationFirst, all unprocessed measurement spectra were dark-corrected by subtracting the detector dark noise spectrum, , which was registered immediately after each measurement series. The dark-corrected spectra were then filtered and resampled in three preprocessing steps: first, the spectrum was equidistantly up-sampled to a resolution of ; second, high-frequency noise and aliasing effects were removed using a phase-invariant low-pass filter (Butterworth of order 4) with a normalized cutoff frequency of 0.02; and third, the spectra was down-sampled to a resolution of and limited to the wavelength region . Visual inspection of the residuals when comparing unfiltered and filtered data displayed only stochastic noise with no trends or patterns. To eliminate influences from changes in lamp intensity and color, all preprocessed measurement spectra were normalized by simultaneously recorded and preprocessed lamp spectra. The unprocessed white reference measurements were identically preprocessed and normalized, and in addition, averaged over time in order to obtain a single white reference spectrum. Last, the lamp-normalized measurement spectra were white-corrected by dividing with the lamp-normalized time-averaged white spectra. The complete normalization process is described in Eq. 1: where denotes averaging over time. The preprocessed and normalized spectrum was used in the analysis of myocardial tissue status.The method used for calibration was originally described by Jacques21 and has, with some modifications, been used by other authors.22, 23 The details are described in Häggblad, 22 and only a brief overview is given here. The method utilizes a modified Beer-Lambert model that takes both absorption and scattering into account when describing the light transport, , in tissue. The expression is given in Eq. 2: where , and , with to being constants determined in a calibration procedure involving measurements on liquid optical phantoms based on milk (scatterer), ink (absorber), and distilled water. The optical properties of the phantom grid were , . The anisotropy factor, , of milk at was set to 0.84 according to a previous study,24 resulting in . The calibrated light transport model was denoted .2.5.Tissue Chromophore Selection and Spectral AnalysisThe major chromophores in myocardial tissue are oxygenized and deoxygenized hemoglobin and myoglobin ( , Hb, , and Mb, respectively), water (W), and fat (lipid).7, 14, 22 In addition to the major chromophores described earlier, it has been shown that methemoglobin (metHb) and changes in oxidation status of cyt aa3 in the myocardium can be detected by DRS.10, 18, 25 To investigate whether inclusion of these additional chromophores resulted in a substantially improved model, two different setups were evaluated: one with only the major chromophores (Hb, , Mb, , and W) included in the absorption model (model I); and one including, in addition to the major chromophores, metHb and the oxidized (cyt aa3,ox) and reduced (cyt aa3,red) form of cyt aa3 (model II). In this study, lipid absorption was not included in the light transport model because of its very low intramuscular concentration26, 27 compared to the epicardial fat layer.22 According to several studies, the water content was set to 75% (Refs. 21, 22, 28). Because of the very similar absorption spectra of hemoglobin and myoglobin (Mb is red-shifted compared to Hb), only the sum of the tissue fractions of these two chromophores was determined. However, to account for this shift, hemoglobin absorption spectra, with an adaptive wavelength transition of , was used in our absorption model [Eq. 3]. The adaptive transition was implemented as a fitting parameter when solving the inverse problem. Equation 3 shows the expression for the total myocardial tissue absorption for model II. The expression for model I is obtained by forcing and to be equal to zero. In Eq. 3, is the tissue fraction, is the oxygen saturation, is the cyt aa3 tissue fraction, is the oxidation status of cyt aa3, and is the metHb tissue fraction. It should be noted that absorption data is often provided in terms of the specific extinction coefficient, . The conversion from to can be found in Sec. 6.The reduced scattering coefficient was modeled according to a previous study by Häggblad22 : where and are the fitting parameters.By combining and Eq. 3, expressions for light transport in myocardial tissue are obtained for both models and are denoted and , respectively. The unknown parameters in Eqs. 3, 4 ( , , , , , , and ) were determined by minimizing the expressions and in a least-squares sense using the Levenberg-Marquardt algorithm. The Levenberg-Marquardt algorithm has been shown useful when solving nonlinear systems in a least-squares sense.29 The model spectra giving the optimum solutions were denoted and , respectively. For both models, visual inspection of was performed, and the goodness of fit was evaluated by calculating the Pearson product-moment correlation coefficient . Visual inspection of is a very intuitive way of identifying wavelength interval where the model fails to fit the measured data. In turn, this reveals information about possible missing chromophores in the light transport model. The absorption data of hemoglobin and water were compiled from Zijlstra and Buiteveld, 30, 31 respectively. The absorption data for cyt aa3,ox and cyt aa3,red were compiled from the BORL home page,19 and absorption data for metHb was compiled from Zijlstra 30 As the cyt aa3 absorption data were not available for wavelengths shorter than (Ref. 19), while the sensitivity of the spectrometer was considered too poor above , the spectral analysis was limited to the wavelength region . In group A, spectra from the last minute of each level were averaged over time, and the curve-fitting was applied on the resulting spectra. In group B, spectra from the last of each level and LVAD speed were averaged over time, and the curve-fitting was applied on the resulting spectra. 2.6.Statistical AnalysisSummarized data from groups A and B are calculated from fitting an average spectrum over the recording interval. The effect of on (groups A and B) was evaluated on data from fitting individual spectra using an ANOVA repeated measures design, with calves and as factors. The effect of LVAD pump speed on (calf B1) was evaluated on data from fitting individual spectra using an ANOVA repeated measures design, with subject and LVAD pump speed as factors. 3.Results3.1.Spectral FittingTo illustrate the effect of adding cyt aa3 and metHb as chromophores in the light transport model, two spectra with varying were selected. The curve-fits for model I (dashed black), model II (solid black), and the measured spectrum (solid gray) are shown in Figs. 2 and 3 . Fig. 3(a) Representative measured spectrum and model spectra ( and ) from calf A4, 21% (gray). (b) . 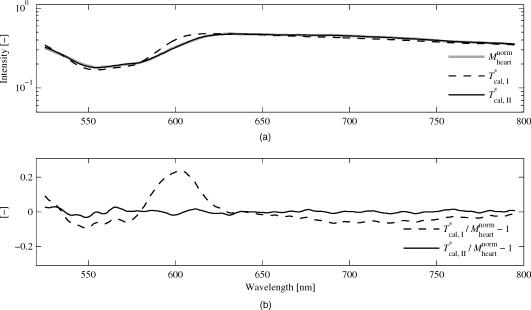 Model I yielded characteristic spectral misfits in the wavelength region . The use of model II resulted in improved spectral fitting, and the characteristic spectral misfits were eliminated. The calculated using model II, compared to from model I, differed only by in the low case (16.7% compared to 17.6%, respectively), and by 2.6% in the high case (37.7% compared to 35.1%, respectively). The corresponding values for differences in were 0.1% in both the high and low case. The -values for the two different curve-fits versus measured spectra from the last minute of phase of group A calves showed that model II resulted in higher -values in all five cases . Taken together, the improved -values and the eliminated spectral misfit characteristics in the residuals supported the choice of model II for further analysis. The wavelength shift in the Hb spectrum was on average in group A , , and during , 50%, and 100%, respectively. In group B, the average wavelength shift was . 3.2.Intramyocardial Light ScatteringThe parameters and in Eq. 4 were averaged for each calf. In group A, ranged from , and was in the range 2.43 to 2.84. Corresponding values for group B were and 2.18 to 3.91 . The average calculated for both groups was at and at , which are both commonly used laser wavelengths in biomedical optics. 3.3.Intramyocardial Tissue ChromophoresThe results for chromophore tissue fractions and oxygenation/oxidation status are presented for each group (A and B) separately. In addition, for group B, results for the two calves are presented as separate cases. 3.3.1.Group A (provocations with changes in )As a representative case, Fig. 4 shows the time-resolved for calf A2 during the variations in . Fig. 4in calf A2 as a function of time during three different and during cardiac arrest at the end of the experiment. The last minute in each phase is marked with vertical dashed lines.  As can be seen in Fig. 4, there is an increase in that follows the changes from 21% via 50% to 100%. Approximately after each change, the reached a stable value, suggesting that the during the last of each level is an accurate interval when calculating average values for each . Cyclic variations in and were detected throughout the measurements for all cases, although with varying magnitude of the variation. As an illustrative example of a case with high variation, for the first of calf A1, is shown in the left panel of Fig. 5 . The right panel illustrates the correlation between and during the same period. Fig. 5(a) as a function of time during the first in calf A1. (b) Relationship between and including linear fitting during the first in calf A1.  The left panel in Fig. 5 shows distinct periodical behavior of . The number of local maxima was determined by visual inspection to 34 during , giving a frequency of . This was compared to the heart rate observed by the ECG monitor, which was . The right panel displays an inverse relationship between and . The -value of the fitted straight line was 0.94. Negative correlations between and were found for four of five calves. Similarly, the relationship between the wavelength shift of the Hb spectrum and the was evaluated in order to determine whether the wavelength shift could be related to Mb during blood volume pulsations. However, the correlation between the wavelength shift and was slightly positive in calves A1 to A3, slightly negative in A4, and without any trend in A5. , , , and average values (over the last minute) of each level are presented in Table 1 . was on average 97%, ranging from 82% to 100%, and displayed no correlations with . Thus, data were omitted from Table 1. Table 1Average chromophore tissue fractions and SHb+Mb for the last minute of every FI,O2 level.
As can be seen in Table 1, calves A1 to A4 responded with an increase in when increased. Increasing 21% to 50% to 100%, gave average values for , , and , respectively. Hence, the average increases in were 4.6% and 4.5% when was increased from 21% to 50% and from 50% to 100%, respectively. The effect of on was significant ( , ANOVA). The varied between calves ( , ANOVA), as did the response to ( , ANOVA). Calves A1 to A3 had a similar step-wise increase in their , while for A4, a larger response was seen when increased from 50% to 100% than from 21% to 50%. For A5, decreased when increased from 50% to 100%. 3.3.2.Group B (provocations with changes in LVAD speed and )The time-resolved for calf B1 is shown in Fig. 6 . It can be seen in Fig. 6 that the did not reach stable levels during the provocations . For this reason, spectra from the 10 last seconds for every level were averaged when calculating the tissue fractions and and levels of the chromophores. was on average 98%, ranging from 84% to 100% throughout measurements in calf B1, and displayed no correlations with . The time-resolved for calf B2 is shown in Fig. 7 . Fig. 6as a function of time during variations in LVAD speed and in calf B1. Note: Last in each phase are marked with vertical dashed lines.  Fig. 7as a function of time during variations in during constant LVAD speed in calf B2. Note: Last in each phase are marked with vertical dashed lines. 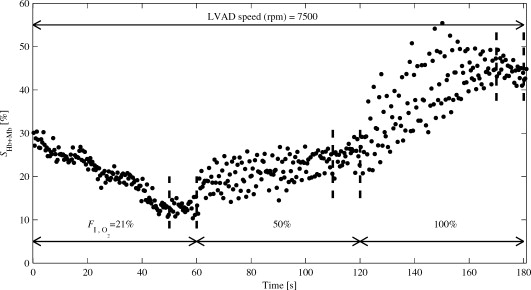 A similar oxygenation trend as in group A was observed during the variations. It should be noted that during the first level (21%; ), did not reach a stable level. As in calf B1, spectra from the last were averaged when calculating chromophore tissue fractions and and levels. The chromophore tissue fractions and oxygenation levels for both calves are summarized in Table 2 . was on average 96%, ranging from 85% to 100%, throughout the measurements in calf B2, and are omitted in Table 2. Table 2Average chromophore tissue fractions and SHb+Mb for the last 10s of every phase.
In calf B1, only a small increase from 18.8% to 20.7% was found in when increasing the LVAD speed from to . When increasing the LVAD speed from to , there was a larger increase, from 20.7% to 38.4%. The increased with LVAD pump speed ( , ANOVA). During the following phases with constant LVAD speed and variations in 21%, , 50% , 100% , increased significantly ( , ANOVA). 4.DiscussionWe have presented a calibrated diffuse reflectance spectroscopy method for in vivo determination of myocardial tissue light scattering properties and chromophore tissue fractions, as well as the sum of hemoglobin and myoglobin oxygenation and cytochrome oxidation. A custom-made fiber-optic probe was designed to enable intramyocardial probe placement. The method relies on an empirical light transport model together with calibration measurements on liquid optical phantoms,21 which previously has been found applicable for in vivo determination of myocardial oxygenation.22 The method was evaluated during hemodynamic and respiratory provocations in seven calves, enrolled in two different studies. 4.1.Spectral FittingIn our previous study of myocardial oxygenation estimation with DRS,22 we found characteristic spectral misfits in model adaption to measured spectra. For model I, the lowest value for the model adaption to measured spectra from group A, , was 0.954. The corresponding value for the model II was . Averaged over calves A1 to A5, the value improved from 0.965 (model I) to (model II). When using model II, the residual spectra, defined as , displayed no systematic trends as function of wavelength. Furthermore, the introduction of a fitting parameter in the model allowing wavelength shifting (up to ) of hemoglobin absorption spectra, naturally contributed to the improved spectra fit compared to our previous study. When including additional chromophores, it is a necessity that their absorption spectra are unique for the chosen wavelength region. This means that none of the chromophores should have an absorption spectrum that could be expressed as a linear combination of other included chromophores. In this study, where the wavelength region was used, all included chromophores displayed unique absorption spectra. 4.2.Intramyocardial Light ScatteringThe estimated average value for at of in this study is comparable to that in pig hearts by Gandjbakhche 32 ( at )32 Reported values by Häggblad are in the range in wavelength region .22 4.3.Intramyocardial Tissue Chromophores4.3.1.Hemo- and myoglobinThe values reported in Table 1 and Table 2 varied from 0.5% to 1.6% and displayed very small and nonsystematic intraindividual variations during changes in and/or LVAD speed. The magnitude of pulsations varied between cases, suggesting that large variations may indicate the presence of a large blood vessel, while smaller variations may indicate a probe placement in an area dominated by capillaries. It should be noted that the magnitude of the variations in may be affected by movement artifacts due to myocardial contractions, although it is fixated by epicardial suturing. Due to the contractile movement of the myocardium, it is likely that the pressure between the probe tip and the surrounding tissue varies. At such high pressure (contractile phase), Hb molecules in front of the probe may be forced away, causing to drop. When analyzing individual spectra, cyclic variations in with identical frequency as the heart rate were observed. Since myoglobin is bound to muscle tissue,33 it is reasonable to assume that variations in should be associated with changes mostly in hemoglobin tissue fraction. Moreover, myoglobin is nearly fully oxygenated at normal working conditions,18 and releases oxygen only at very low partial oxygen pressure,34 which is not the case during the conditions in this study. Our observed variations in should therefore be interpreted as a change mostly in hemoglobin oxygenation. The relationship between the pulsatile variations in the wavelength shift of the Hb spectrum and the did not support the idea that the relative Hb and Mb contributions can be determined from the wavelength shift. However, the red-shift of Mb is small (about ), which demands very high wavelength accuracy in the recordings and also suggests that reference spectra obtained with the same spectrometer should be used. We chose not to use more sophisticated methods for separation of hemoglobin and myoglobin oxygenation, despite the fact that other authors14, 35, 36 have made successful attempts to distinguish between hemoglobin and myoglobin absorption influence on both in vitro and in vivo diffuse reflectance spectra recordings. In the majority of cases, we observed a strong negative correlation between and If the assumptions of constant myoglobin tissue fraction and saturation (nearly 100%) holds true, a likely explanation for the negative versus correlation is that when decreases, the relative influence of myoglobin absorption in the measured spectra increases, and since myoglobin is nearly fully oxygenated, will increase. However, maximum values of 33.3% (group A; A1) and 44.3% (group B; B2), respectively, suggest that the hemoglobin absorption contribution is predominant throughout the measurements. In group A, 4 of 5 calves responded with increased values when inspired was increased. During ventilation with room air , the average was 20.0%, which corresponds to an myocardial oxygen uptake of 80%, which is in agreement with findings from several other studies.37, 38, 39 In calf B1, a small increase in was found when increasing LVAD speed from . When the LVAD speed was further increased to , drastically increased. During all three LVAD speed settings, the was held at 100%. For the following changes in levels during constant LVAD speed, the displayed the same trend as in calves A1 to A4. When increasing in calf B2, increased from 12.9% to 44.3% . The finding that increased with LVAD pump speed was expected, since the purpose of the LVAD is to lower the amount of work needed (and hence lower the oxygen consumption) in the left ventricle, to maintain sufficient systemic and coronary circulation. 4.3.2.Cyt aa3 and metHbThe values reported in Table 1 and Table 2 varied from 1.2% to 2.0% and displayed very small and nonsystematic intraindividual variations during changes in and/or LVAD speed. By assuming a molecular weight of for cyt aa3 (Ref. 40), the findings for gives a cyt aa3 tissue concentration ranging from tissue wet weight. Reported values for porcine heart in vitro are tissue wet weight.41 The findings of above 96% in both group A and group B, despite low are compatible with findings in pig hearts during LAD occlusion.17 In Parson’s study, decreased only when LAD flow was reduced by more than 40% of resting flow.17 The nonrestricted coronary flow and normoxic conditions present throughout the measurements did not, as expected, display any reduction of the . The obtained fractions of methemoglobin were in the interval 0 to 0.2%, which is in agreement with values reported by Wright 42 5.ConclusionsBy using a calibrated intramuscular probe, the tissue chromophore fractions and the oxygenation and cyt aa3 oxidation status of the myocardial chromophores during respiratory and hemodynamic provocations can be monitored. The presented DRS analysis algorithm is capable of delivering results that are physiologically reasonable and in agreement with others. Model inclusion of the chromophores metHb, cyt aa3, ox, and cyt aa3, red, in addition to Hb, , and W, resulted in nonsystematic residual spectra. The presented diagnostic tool follows the way of the oxygen from hemo- and myoglobin to the last step of the oxidative phosphorylation and has the potential for enabling early detection of tissue hypoxia in the heart. However, future studies should include coronary flow provocations and pharmaceutically induced alterations of the mitochondrial ability to utilize oxygen in the ATP formation process. Furthermore, in these future studies, it would be advisable to include, in addition to the presented DRS technique, additional measurement techniques such as microdialysis for cellular metabolism analysis43 and intermittent coronary sinus blood samples for global in vitro analysis of hemoglobin oxygenation. AcknowledgmentsThis study was financed by VINNOVA and Perimed AB through the SAMBIO research collaboration program between companies and academia within bioscience (VINNOVA, D. no. 2008-00149), by the Swedish Research Council (D. no. 2005-3934), and by NovaMedTech (supported by the European union Regional Development Fund). The authors would also like to thank the staff at the Laboratory for Experimental Animal Studies, Faculty of Health Sciences, Linköping University. ReferencesR. I. Williams,
R. Haaverstad,
G. Sianos,
E. Vourvouri, and
A. G. Fraser,
“Perioperative tissue Doppler echocardiography and bypass graft flowmetry in patients undergoing coronary revascularization: predictive power for late recovery of regional myocardial function,”
J. Am. Soc. Echocardiogr, 15
(10, II), 1202
–1210
(2002). https://doi.org/10.1067/mje.2002.122965 0894-7317 Google Scholar
R. Haaverstad,
N. Vitale,
O. Tjomsland,
A. Tromsdal,
H. Torp, and
S. O. Samstad,
“Intraoperative color Doppler ultrasound assessment of LIMA-to-LAD anastomoses in off-pump coronary artery bypass grafting,”
Ann. Thorac. Surg., 74
(4), 1390
–1394
(2002). https://doi.org/10.1016/S0003-4975(02)04058-4 0003-4975 Google Scholar
L. Lovstakken,
K. S. Ibrahim,
N. Vitale,
S. T. Henriksen,
I. Kirkeby-Garstad,
H. Torp, and
R. Haaverstad,
“Blood flow imaging: a new two-dimensional ultrasound modality for enhanced intraoperative visualization of blood flow patterns in coronary anastomoses,”
J. Am. Soc. Echocardiogr, 21
(8), 969
–975
(2008). https://doi.org/10.1016/j.echo.2008.01.019 0894-7317 Google Scholar
W. J. Parsons,
J. C. Rembert,
R. P. Bauman,
J. C. Greenfield Jr., C. A. Piantadosi,
“Dynamic mechanisms of cardiac oxygenation during brief ischemia and reperfusion,”
Am. J. Physiol. Heart Circ. Physiol., 259
(5), 1477
–1485
(1990). 0363-6135 Google Scholar
M. K. Al-Obaidi,
P. J. Etherington,
D. J. Barron,
C. P. Winlove, and
J. R. Pepper,
“Myocardial tissue oxygen supply and utilization during coronary artery bypass surgery: evidence of microvascular no-reflow,”
Clin. Sci., 98
(3), 321
–328
(2000). https://doi.org/10.1042/CS19990138 0323-5084 Google Scholar
Y. Chung,
“Oxygen reperfusion is limited in the postischemic hypertrophic myocardium,”
Am. J. Physiol. Heart Circ. Physiol., 290
(5), H2075
–H2084
(2006). https://doi.org/10.1152/ajpheart.00619.2005 0363-6135 Google Scholar
F. W. Heineman,
V. V. Kupriyanov,
R. Marshall,
T. A. Fralix, and
R. S. Balaban,
“Myocardial oxygenation in the isolated working rabbit heart as a function of work,”
Am. J. Physiol. Heart Circ. Physiol., 262
(1), 255
–267
(1992). 0363-6135 Google Scholar
J. R. Leisey,
D. A. Scott,
L. W. Grotyohann, and
R. C. Scaduto Jr.,
“Quantitation of myoglobin saturation in the perfused heart using myoglobin as an optical inner filter,”
Am. J. Physiol. Heart Circ. Physiol., 267
(2), 645
–653
(1994). 0363-6135 Google Scholar
J. R. Zijp and
J. J. Ten Bosch,
“Optical properties of bovine muscle tissue in vitro: a comparison of methods,”
Phys. Med. Biol., 43
(10), 3065
–3081
(1998). https://doi.org/10.1088/0031-9155/43/10/026 0031-9155 Google Scholar
K. A. Schenkman,
“Cytochrome c oxidation determined by multiwavelength optical spectral analysis in the isolated perfused heart,”
Proc. SPIE, 4613 278
–285
(2002). https://doi.org/10.1117/12.465256 0277-786X Google Scholar
C. Ribacka,
“Redox-linked proton transfer by cytochrome c oxidase,”
64 University of Helsinki,
(2007). Google Scholar
L. H. Opie, Heart Physiology from Cell to Circulation, 4th ed.Lippincott, Williams & Wilkins, Philadelphia, PA
(2004). Google Scholar
C. E. Cooper and
R. Springett,
“Measurement of cytochrome oxidase and mitochondrial energetics by near-infrared spectroscopy,”
Philos. Trans. R. Soc. London, Ser. B, 352
(1354), 669
–676
(1997). https://doi.org/10.1098/rstb.1997.0048 0962-8436 Google Scholar
K. A. Schenkman,
D. R. Marble,
E. O. Feigl, and
D. H. Burns,
“Near-infrared spectroscopic measurement of myoglobin oxygen saturation in the presence of hemoglobin using partial least-squares analysis,”
Appl. Spectrosc., 53
(3), 325
–331
(1999). https://doi.org/10.1366/0003702991946532 0003-7028 Google Scholar
A. Mayevsky and
B. Chance,
“Oxidation-reduction states of NADH in vivo: from animals to clinical use,”
Mitochondrion, 7
(5), 330
–339
(2007). https://doi.org/10.1016/j.mito.2007.05.001 1567-7249 Google Scholar
F. F. Jöbsis,
J. H. Keizer,
J. C. LaManna, and
M. Rosenthal,
“Reflectance spectrophotometry of cytochrome aa3 in vivo,”
J. Appl. Physiol.: Respir., Environ. Exercise Physiol., 43
(5), 858
–872
(1977). 0161-7567 Google Scholar
W. J. Parsons,
J. C. Rembert,
R. P. Bauman,
F. G. Duhaylongsod,
J. C. Greenfield Jr., C. A. Piantadosi,
“Myocardial oxygenation in dogs during partial and complete coronary artery occlusion,”
Circ. Res., 73
(3), 458
–464
(1993). 0009-7330 Google Scholar
A. E. Arai,
C. E. Kasserra,
P. R. Territo,
A. H. Gandjbakhche, and
R. S. Balaban,
“Myocardial oxygenation in vivo: optical spectroscopy of cytoplasmic myoglobin and mitochondrial cytochromes,”
Am. J. Physiol. Heart Circ. Physiol., 277
(2), 683
–697
(1999). 0363-6135 Google Scholar
“BORL Tissue Spectra,”
(2009) http:www.medphys.ucl.ac.uk/research/borl/research/NIR_topics/spectra/spectra.htm Google Scholar
L. Hübbert,
B. Peterzén,
S. Träff,
B. Janerot-Sjöberg, and
H. Ahn,
“Axial flow pump treatment during myocardial depression in calves: an invasive hemodynamic and echocardiographic tissue Doppler study,”
ASAIO J., 54 367
–371
(2008). https://doi.org/10.1097/MAT.0b013e31817fd8de 1058-2916 Google Scholar
S. L. Jacques,
“Optical fiber reflectance spectroscopy,”
(2009) http://omlc.ogi.edu/news/oct03/saratov/index.htm Google Scholar
E. Häggblad,
T. Lindbergh,
M. G. D. Karlsson,
H. Casimir-Ahn,
E. G. Salerud, and
T. Strömberg,
“Myocardial tissue oxygenation estimated with calibrated diffuse reflectance spectroscopy during coronary artery bypass grafting,”
J. Biomed. Opt., 13
(5), 054030
(2008). https://doi.org/10.1117/1.2976433 1083-3668 Google Scholar
P. R. Bargo,
S. A. Prahl,
T. T. Goodell,
R. A. Sleven,
G. Koval,
G. Blair, and
S. L. Jacques,
“In vivo determination of optical properties of normal and tumor tissue with white light reflectance and an empirical light transport model during endoscopy,”
J. Biomed. Opt., 10
(3), 034018
(2005). https://doi.org/10.1117/1.1921907 1083-3668 Google Scholar
T. Lindbergh,
I. Fredriksson,
M. Larsson, and
T. Strömberg,
“Spectral determination of a two-parametric phase function for polydispersive scattering media,”
Opt. Express, 17
(3), 1610
–1621
(2009). https://doi.org/10.1364/OE.17.001610 1094-4087 Google Scholar
J. Lee,
N. El-Abaddi,
A. Duke,
A. E. Cerussi,
M. Brenner, and
B. J. Tromberg,
“Noninvasive in vivo monitoring of methemoglobin formation and reduction with broadband diffuse optical spectroscopy,”
J. Appl. Phys., 100
(2), 615
–622
(2006). https://doi.org/10.1152/japplphysiol.00424.2004 0021-8979 Google Scholar
S. Fleischer,
G. Rouser,
B. Fleischer,
A. Casu, and
G. Kritchevsky,
“Lipid composition of mitochondria from bovine heart, liver, and kidney,”
J. Lipid Res., 8
(3), 170
–180
(1967). 0022-2275 Google Scholar
D. Y. Fei and
K. K. Shung,
“Ultrasonic backscatter from bovine tissues,”
IEEE Ultrasonics Symposium, 677
–680 IEEE, Piscataway, NJ
(1984). Google Scholar
B. A. Wittenberg,
J. B. Wittenberg, and
I. R. Katz,
“Oxygen transport in isolated cardiac myocytes,”
Basic Res. Cardiol., 80 75
–77
(1985). 0300-8428 Google Scholar
J. Nocedal and
S. J. Wright,
“Algorithms for nonlinear least-squares problems,”
Numerical Optimization, 258
–262 Springer Science and Business Media, New York (2008). Google Scholar
W. G. Zijlstra,
A. Buursma, and
O. W. van Assendelft,
“Definitions and terminology,”
Visible and Near Infrared Absorption Spectra of Human and Animal Haemoglobin–Determination and Application, 9
–16 VSP, Ridderkerk, The Netherlands
(2000). Google Scholar
H. Buiteveld,
J. M. H. Hakvoort, and
M. Donze,
“The optical properties of pure water,”
Proc. SPIE, 2258 174
–183
(1994). https://doi.org/10.1117/12.190060 0277-786X Google Scholar
A. H. Gandjbakhche,
R. F. Bonner,
A. E. Arai, and
R. S. Balaban,
“Visible-light photon migration through myocardium in vivo,”
Am. J. Physiol. Heart Circ. Physiol., 277
(2), 698
–704
(1999). 0363-6135 Google Scholar
G. J. Tortora and
S. Reynolds Grabowski,
“The cardiovascular system: Blood vessels and hemodynamics,”
Principles of Anatomy and Physiology, 696
–756 John Wiley & Sons, Inc., Hoboken, NJ
(2003). Google Scholar
K. A. Schenkman,
D. R. Marble,
D. H. Burns, and
E. O. Feigl,
“Myoglobin oxygen dissociation by multiwavelength spectroscopy,”
J. Appl. Physiol., 82
(1), 86
–92
(1997). 8750-7587 Google Scholar
K. A. Schenkman,
D. R. Marble,
D. H. Burns, and
E. O. Feigl,
“Optical spectroscopic method for in vivo measurement of cardiac myoglobin oxygen saturation,”
Appl. Spectrosc., 53
(3), 332
–338
(1999). https://doi.org/10.1366/0003702991946541 0003-7028 Google Scholar
L. S. L. Arakaki,
M. J. Kushmerick, and
D. H. Burns,
“Myoglobin oxygen saturation measured independently of hemoglobin in scattering media by optical reflectance spectroscopy,”
Appl. Spectrosc., 50
(6), 697
–707
(1996). https://doi.org/10.1366/0003702963905763 0003-7028 Google Scholar
B. I. Levy,
E. Pinard, and
J.-B. Michel,
“Transmural gradient of tissue gas tensions in the canine left ventricular myocardium during coronary clamping and reactive hyperemia,”
Pflugers Archiv. Eur. J. Physiol., 407
(4), 388
–395
(1986). https://doi.org/10.1007/BF00652623 Google Scholar
J. Vinten-Johansen and
H. R. Weiss,
“Oxygen consumption in subepicardial and subendocardial regions of the canine left ventricle. The effect of experimental acute valvular aortic stenosis,”
Circ. Res., 46
(1), 139
–145
(1980). 0009-7330 Google Scholar
T. M. Nosek, Essentials of Human Physiology, http://imc.meded.com/integrated/demos/hpdemo/program/start.htm Google Scholar
S. Ferguson-Miller,
“Mammalian cytochrome c oxidase, a molecular monster subdued,”
Science, 272
(5265), 1125
(1996). https://doi.org/10.1126/science.272.5265.1125 0036-8075 Google Scholar
R. S. Balaban,
V. K. Mootha, and
A. Arai,
“Spectroscopic determination of cytochrome c oxidase content in tissues containing myoglobin or hemoglobin,”
Anal. Biochem., 237
(2), 274
–278
(1996). https://doi.org/10.1006/abio.1996.0239 0003-2697 Google Scholar
R. O. Wright,
W. J. Lewander, and
A. D. Woolf,
“Methemoglobinemia: etiology, pharmacology, and clinical management,”
Ann. Emerg. Med., 34
(5), 646
–656
(1999). https://doi.org/10.1016/S0196-0644(99)70167-8 0196-0644 Google Scholar
J. Pöling,
A. Leptien,
S. Klaus,
W. Rees,
E. Kraatz,
K. Wiebe, and
L. Bahlmann,
“Analysis of the myocardial metabolism by microdialysis during open beating heart surgery,”
Scand. Cardiovasc. J., 41
(2), 114
–119
(2007). https://doi.org/10.1080/14017430601091706 1401-7431 Google Scholar
S. A. Prahl,
“Optical absorption of hemoglobin,”
(2009) http://omlc.ogi.edu/spectra/hemoglobin/index.html Google Scholar
AppendicesAppendixThe absorption data for all chromophores are given in specific extinction coefficients . To be included in the light transport model, has to be translated into the absorption coefficient . A general description of this conversion has been described by Prahl44 and is shown in Eq. 5; where is the substance density , and is the molecular weight of the specific substance.For the hemoglobin protein, consisting of four heme units, the molecular weight was set to . For cyt aa3, where each functional unit consists of 13 subunit enzymes containing haem a, haem a3, CuA, and CuB, the molecular weight was set to . To convert the specified tissue fractions to a molecular amount (e.g., mol per gram tissue), simply divide the fraction by the corresponding molecular weight. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||