
|
|
1.IntroductionPreviously, we have shown that a 200-fs near-infrared (NIR) laser induced focal point–specific damage in the genomic deoxyribose nucleic acid (DNA) of live cells, and such damage led to the activation of the DNA damage response (DDR).1 Further studies on fully condensed chromosomes during cell division (mitosis) demonstrated the activation of the DDR when NIR laser focal-point damage was produced on chromosome arms in mitotic (Potorous tridactylus) cells.2 When a second harmonic green ps Nd:YVO4 laser was used to expose the ends (tips) of chromosomes as they separated during mitotic anaphase, a significant percent of cells either delayed cytokinesis or did not divide at all. This occurred even when a single chromosome tip was damaged.3 The present study was designed to determine if the damage to a single chromosome end, which is known to contain the telomere, activates the DDR and repair pathways. We also need to understand whether laser-induced microirradiation at different chromosomal locations leads to distinct recruitment of DDR and repair factors. Analyzing the recruitment of factors at different chromosomal locations will determine if laser microirradiation, particularly with the commonly used NIR fs lasers, can be used to study the activation of DDR and repair pathways regardless of the chromosome location and as well, shed light on the functionality of the telomere during mitosis. It is well established that telomeres occur at the ends of chromosomes. They are nucleoprotein structures composed of DNA double-stranded 5′-TTAGGG-3′ repeats and a single-stranded G-rich 3′ overhang.4 The ends of linear chromosomes structurally resemble double-stranded DNA breaks (DSBs) but are protected by a nucleoprotein structure called shelterin.4–6 Interphase mammalian telomeres can be “uncapped” by the loss of function of shelterin components, which result in the activation of the DDR.4 Although there have been studies on the activation of DDR at uncapped telomeres in the interphase stages of the cell cycle, there are no studies on the activation of DDR at the telomere-containing chromosome ends while the cell is in mitosis, especially during anaphase. Most existing methods used to study mitotic DDR activation and the recruitment of repair proteins at mammalian telomeres [telomere-dysfunction induced foci (TIFs)]7 utilize microtubule depolymerizing drugs, which result in uncapping of chromosome ends leading to long-term mitotic arrest.8 DNA breaks can also be produced in mitotic chromosomes by incubating cells with depolymerizing drugs, but only recruitment of DDR factors to DNA breaks has been observed and not recruitment of repair proteins.9 Recent evidence also shows that DSBs persist at telomeres in interphase and are unable to get repaired, whereas DSBs in chromosomal DNA are efficiently repaired.10 Earlier studies have shown that laser-induced DNA damage results in the activation of the DDR in interphase mammalian cells.11–13 Additionally, based on the observed accumulation of phosphorylated histone H2AX (γH2AX), a well-known DSB marker, it has been shown that NIR lasers without the use of sensitizing agents are capable of producing DSBs on a submicrometer scale without unwanted structural damage to the nucleus of living cells.14–19 Our goal in this study is to determine if NIR laser-induced DNA breaks at telomere-containing chromosome ends in mitotic anaphase results in the activation of the DDR and recruitment of repair proteins. An additional goal is to determine if this correlates with the delay in mitotic cytokinesis reported previously,3 and, in particular, whether there is a difference in the DDR factors recruited to DNA breaks when a single chromosome end is damaged as opposed to a chromosome arm. The flat morphology, small number, and large size of chromosomes () as well as the close sequence identity of these cells with those of humans, mice, and rats (80% to 90%) make this cell type ideal to study the DDR processes using selective short-pulse NIR laser microirradiation.20–22 2.Materials and Methods2.1.Cell Line and Cell CulturePotorous tridactylus (-male) kidney epithelial cells (American Type Culture Collection ATCC, CCL 56) were grown in Gibco advanced minimum essential medium (Invitrogen) supplemented with l-glutamine, 4% fetal bovine serum, and antibiotics. Cells were incubated at 37°C with 5% .3 Three days before experiments, cells were trypsinized (TrypLE TM Express, Life Technologies) and plated on 35-mm gridded imaging dishes (MatTek) at . 2.2.Laser Exposure and DosimetryThe custom RoboLase ablation system uses a 200-fs, 76-MHz Ti:Sapphire NIR 800-nm laser (Coherent Inc., Santa Clara, CA) coupled to a motorized inverted Zeiss microscope (Axiovert 200 M) with a 37°C, 35-mm culture dish stage (Warner Instruments, LLC, Connecticut, USA). Custom LabView software was developed for the use of the automated microscope system and laser.23 Single telomere-containing chromosome ends and chromosome arms of live anaphase cells were irradiated with a diffraction-limited (0.7-μm diameter) focal spot.2 The laser was focused by front-surfaced mirrors to a Zeiss 63×/1.4 NA phase contrast oil objective. To determine the irradiance at the focal spot, the transmission of the objective at 800 nm was measured using the double-objective method.2 Physical measurement of the beam power was made prior to beam entry into the back aperture of the microscope objective using a FieldMaxII-TOP power laser meter (Coherent Inc., Santa Clara, CA). Based upon the measurement of the entry power into the objective, the objective transmission, and the estimated focal spot size, the laser irradiance at the focal point was determined to be . To study the recruitment of DDR and repair proteins at microirradiated focal spots, individual microirradiated anaphase cells were monitored after laser microirradiation (minutes) and fixed for subsequent fluorescence in situ hybridization (FISH), terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL), or antibody staining. 2.3.FISH and TUNEL Labeling AssaysMicroirradiated single chromosome ends and chromosome arms of individual cells cultured on gridded dishes were fixed after laser focal-point microirradiation with 3.7% formaldehyde in Tris-buffered saline (TBS) for 10 min at room temperature (RT). Dishes were washed three times with phosphate-buffered saline (PBS) and were left at 4°C overnight. Telomeres were visualized using FISH with a Cy3-congugated (TTAGGG)-PNA probe as described in the manufacturer’s instructions (DAKO, Carpinteria, CA). Cells were later permeabilized twice with PBS/0.2% Triton X-100 for 10 min at RT and then washed three times in PBS-ethylenediamine tetra-acetic acid (EDTA) for 5 min followed by one wash with PBS. To label DNA breaks on laser microirradiated samples, cells were incubated with enzyme/label solution mix (TUNEL, Roche) in a humidified chamber at 37°C for 1 h. After the reaction, cells were washed three times on a shaker in PBS-EDTA for 5 min to reduce background staining. Samples were visualized and images acquired using a objective on a Zeiss inverted microscope (Axiovert 200 M) equipped with a Hamamatsu Orca CCD camera. Images were analyzed using ImageJ software (NIH, Bethesda, MD). 2.4.Immunofluorescence and ImagingTo observe the recruitment of DDR factors and repair proteins at single microirradiated chromosome ends or internal chromosome arms, cells grown on gridded culture dishes were fixed with 3.0% formaldehyde TBS for 10 min at RT and placed on ice. Cells were permeabilized with 0.5% Triton X-100 for 10 min at RT, washed twice with PBS for 5 min at RT, and incubated with blocking solution (10% calf serum, 1% BSA/PBS) for 1 h at RT. Cells, subsequently, were washed once in PBS for 5 min at RT and then stained with a primary antibody solution of 3% BSA/PBS overnight at 4°C. The following primary antibodies were used: anti-γ-H2AX (07-164; Millipore), anti-Nbs1 (NB100-143, Novus Biological), phospho-Chk1Ser345 (2348, Cell Signaling), phospho-Chk2Thr68 (2661, Cell Signaling), anti-PCNA (2586, Cell Signaling), phospho-p53Ser15 (sc-101762, Santa Cruz Biotechnlogy, Inc.), anti- (sc-71471, Santa Cruz Biotechnology, Inc.), and anti-Rad51 (sc-53428, Santa Cruz Biotechnology, Inc.). After antibody incubation, cells were washed twice in PBS/0.05% Tween 20 for 5 min at RT and incubated with secondary antibodies (Invitrogen; ) for 1 h at RT. Cells were washed twice with PBS/0.05% Tween 20 for 5 min at RT and the DNA was stained with 4,6-diamidino-2-phenylindole ( in PBS) for 5 min at RT. Samples were imaged as described in the previous section. 3.Results3.1.Laser Microirradiation to Single ChromosomesWe examined the DDR proteins recruited to either a single chromosome end or a chromosome arm (distant from the telomere-containing end) during anaphase onset after DNA breaks were produced with the NIR laser.2,12 Two methods were used to verify that the telomere of a single anaphase chromosome end was damaged: (1) FISH using a Cy3-conjugated (TTAGGG)-PNA probe against telomeric DNA and (2) TUNEL to visualize DNA breaks [Fig. 1(a)]. The results demonstrate that the DNA breaks can be induced by focal point laser NIR microirradiation at either chromosome ends or chromosome arms. A previous study has shown that the laser microirradiation of mitotic chromosome DNA results in “paling” at the site of damage followed by the gradual formation of phase-dark material, demonstrated to be the result of the accumulation of DDR factors.2 To verify that we were obtaining the same response at chromosome ends, either individual chromosome ends or chromosome arms were microirradiated at anaphase and monitored by phase-contrast microscopy for several minutes until the presence of phase-dark material was detected. Following laser microirradiation (10-s postlaser), phase paling was evident at microirradiated chromosome ends and microirradiated internal chromosome arms [Figs. 2(a)(ii) and 2(b)(ii)]. After 120 s, phase-dark material was visible at the damage site corresponding to the accumulation of DSB marker phosphorylated histone H2AX (γH2AX), and the early modification enzyme known to facilitate DNA repair of single-stranded breaks (SSBs): poly(ADP-ribose) polymerase 1 (PARP1) [Figs. 2(a)(iii) and 2(b)(iii)]. Fig. 1Single point laser microirradiation induces localized DNA breaks. Fluorescence in situ hybridization (FISH)/terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) to detect DNA breaks produced by laser microirradiation at a single chromosome end and chromosome arm in anaphase cells. Green, TUNEL staining; red, Cy3-5′-TTAGGG-3′ probe; blue, DAPI to stain DNA. Arrow points to laser-induced DNA breaks. Scale bar 10 im. of independent experiments.  Fig. 2Single point laser-induced DNA breaks activates the DDR. (a) Phase image of cell prior laser microirradiation at telomere-containing chromosome end (prelaser), 10 s after laser microirradiation (postFlaser), and 120-s postlaser. (a′) Inset of (a) of a single chromosome end before microirradiation/prelaser (i), 10-s postlaser (ii), and 120-s postlaser (iii). (b) Phase image of cell prior to laser microirradiation at chromosome arm (prelaser), 10 s after laser microirradiation (postlaser), and 120-s postlaser. (b′) Inset of (b) of a single chromosome arm before microirradiation/prelaser (i), 10-s postlaser (ii), and 120-s postlaser (iii). Cells were fixed at 120 s and stained with anti-γH2AX (green), anti-PARP1 (red), and co-stained with DAPI (blue). Dashed circle shows microirradiated DNA and foci accumulation. Scale bar 10 μm. of independent experiments.  3.2.Activation of DDR at Laser-Induced DNA BreaksRecent work has demonstrated that telomeric damage by long-time mitotic arrest leads to checkpoint activation and cell cycle arrest.8 To determine whether proteins involved in cell cycle arrest are recruited to laser-induced DNA breaks in chromosome ends or chromosome arms at the onset of anaphase, the recruitment of DNA-damage checkpoint kinases Chk1, Chk2, and p53 was assessed by immunofluorescence. Cells were fixed 5-min postlaser exposure either to a chromosome end or to a chromosome arm distant from the end (internal chromosome arm). Immediate recruitment of Chk1 phosphorylation on serine 345 (Ser345) and Chk2 phosphorylation on threonine 68 (Thr68) was observed at damaged chromosome ends and chromosome arms [Figs. 3(a) and 3(b), lanes 1 and 2, ]. In addition, DNA breaks on chromosome ends showed immediate foci accumulation of p53 phosphorylation on serine 15 (Ser15) [Fig. 3(a*), lane 3, ]. Laser-induced DNA breaks at the control internal chromosome arms failed to recruit p53 phosphorylation on serine 15 (Ser15) [Fig. 3(b), lane 3, ]. The recruitment of checkpoint proteins Chk1, Chk2, and p53, which are involved in the DDR (Table 1), confirms the activation of a DDR that is likely responsible for the previously observed delay in cytokinesis.3 Fig. 3Recruitment of checkpoint DDR proteins to single point laser-induced DNA breaks during anaphase onset. (a) Postfixation performed 5 min after chromosome end laser microirradiation (fixed) of anaphase cells. (b) Postfixation performed 5 min after chromosome arm laser microirradiation (fixed). Endogenous DNA damage recognition factors accumulate at laser-induced DNA breaks. DNA breaks are detected with anti-γH2AX; checkpoint response proteins are detected with anti-p-Chk2(Ser345), anti-p-Chk1(Thr68), and anti-p-p53(Ser15). Arrows on fixed cells point to laser-induced DNA breaks. Scale bar 10 μm, of independent experiments.  Table 1Number of cells that show foci accumulation of DNA damage response (DDR) and repair proteins at a single damaged chromosome end or chromosome arm.
3.3.Recruitment of Repair Proteins at Laser-Induced DNA BreaksHaving established the activation of the DDR by inducing DNA breaks at chromosome ends and internal chromosome arms, we next examined whether repair proteins from the two major repair pathways, nonhomologous end joining (NHEJ) and homologous recombination (HR), are able to form foci at laser-induced DNA breaks at specific chromosomal sites. Using an anti- endogenous antibody, induced DNA breaks at chromosome ends and chromosome arms showed foci accumulation of endogenous NHEJ repair . This complex co-localized with DDR sensing factor Nbs1, which served as a control [Figs. 4(a) and 4(b), lane 1, ]. To investigate whether additional factors accumulate at laser-induced DNA breaks at chromosome ends or chromosome arms, we examined the recruitment of PCNA, which is known to be involved in replication and DNA repair.24–26 Endogenous PCNA foci accumulated at damaged chromosome ends and chromosome arms [Figs. 4(a) and 4(b), lane 2, ]. Furthermore, antibody staining for Rad51, a mammalian HR repair factor,27 revealed no detectable fluorescence at localized DNA breaks of anaphase chromosome ends or arms as previously shown in mitotic cells [Fig. 5(a)].28 Fig. 4Recruitment of DNA damage repair factors to single point laser-induced DNA breaks during anaphase onset. (a) Postfixation performed 5 min after chromosome end laser microirradiation (fixed) of anaphase cells. (b) Postfixation performed 5 min after chromosome arm laser microirradiation (fixed). Endogenous DNA damage recognition factors accumulate at laser-induced DNA breaks. DNA breaks are detected with anti-γH2AX, anti-, anti-Nbs1, anti-PCNA antibodies, and counterstained with DAPI (blue). Arrows on fixed cells point to laser-induced DNA breaks. Scale bar 10 μm, of independent experiments. 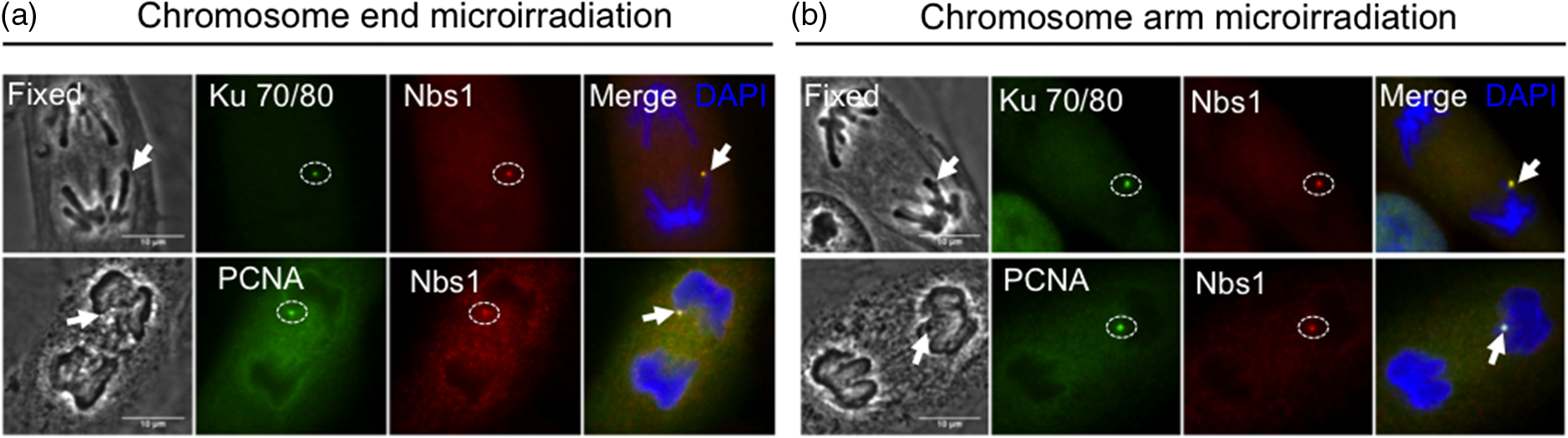 Fig. 5Homologous repair protein Rad51 does not get recruited to anaphase DNA breaks. Postfixation performed 5 min after laser irradiation to a single chromosome end and chromosome arm. Repair proteins are detected with anti-γH2AX (green), anti-Rad51 (red), and co-stained with DAPI (blued). Arrow points to microirradiated chromosome site. Scale bar 10 μm, of independent experiments. 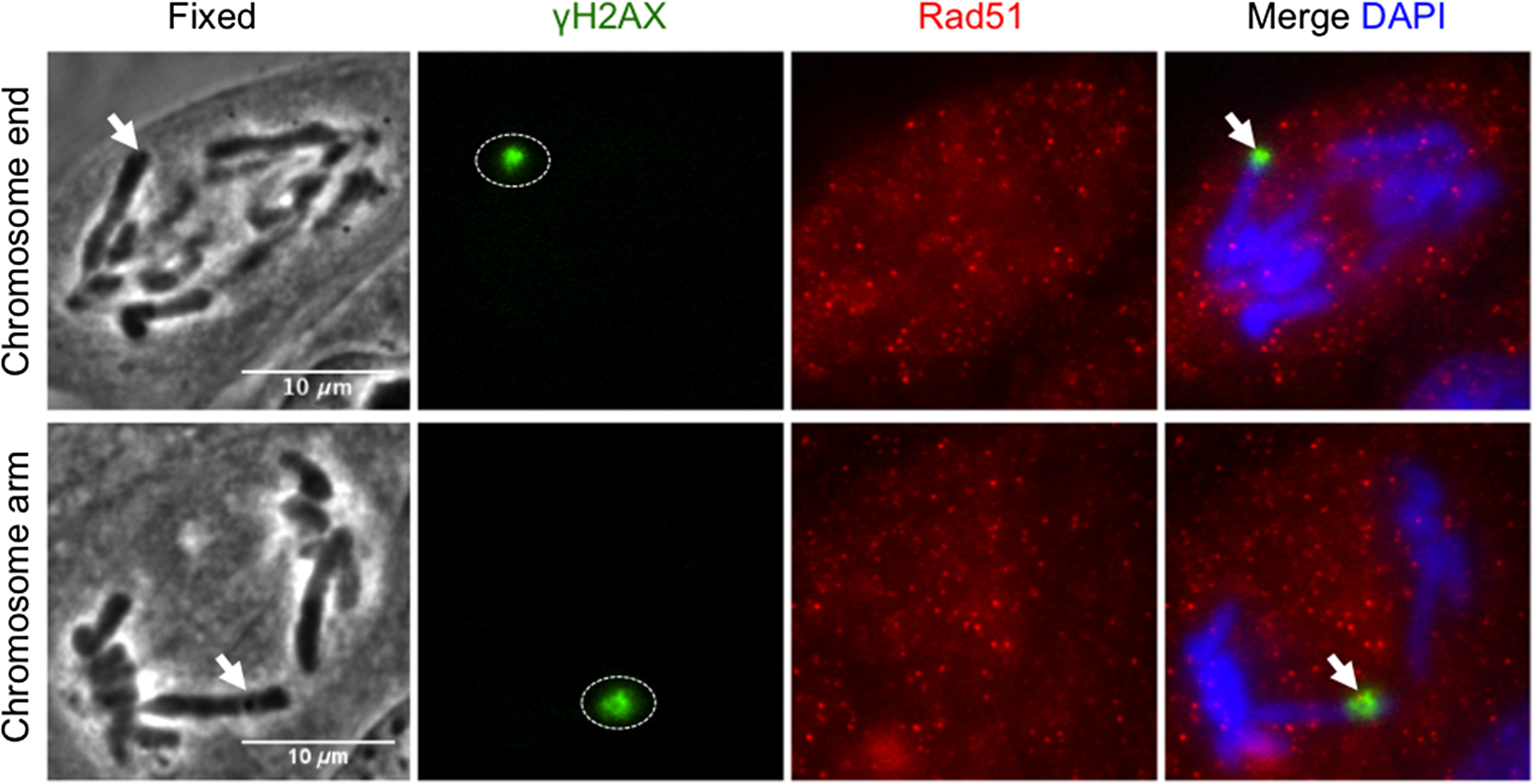 4.DiscussionRecent evidence indicates that chromosome ends of interphase cells lack a repair mechanism compared to the rest of the chromosomes.10 Despite this evidence, few studies have examined the accumulation of DDR factors at damaged chromosome ends during mitosis.8 In fact, there are neither studies nor methods that show the localization of additional DDR factors when the damage is produced while the cell is in mitosis. Furthermore, it remains unclear whether damaged chromosome ends of mitotic cells can activate a full DDR by recruiting proteins from repair pathways such as HR and/or NHEJ. In the present study, DNA breaks at telomere-containing chromosome ends and chromosome arms in vertebrate cells were induced at the onset of anaphase through the use of an NIR femtosecond laser. The irradiance used to produce DNA breaks such as DSBs and SSBs was in the focal spot. At this irradiance, the mechanism of damage was likely a nonlinear multiphoton optical process or possibly a threshold microplasma event localized to the laser focal volume.12 It is unlikely that a significant plasma event occurred because the cell’s survival is indicative of an intact-cell membrane. Because these cells stay relatively flat during mitosis, the outer-cell membrane is within a couple of microns of the focal plane of the laser as shown in the previous transmission electron microscope (TEM) images.2 If a major plasma event occurred at the focal point, the generated shock wave would likely have been sufficient to rupture the outer-cell membrane, resulting in cell death. Previous evidence shows the activation of the DDR by using high irradiance of UV microirradiation at without any plasma formation and using an NIR.12,29 We cannot ignore the damage being produced to the cell using these high irradiances. In our studies, we demonstrate that a lower irradiance, is enough to activate the DDR and repair pathways at chromosome ends and chromosome arms. Additionally, by using TEM it was estimated that the NIR microirradiation produces damage in 10 to 100 Mbp of DNA.2 However, the irradiance used in that study () was slightly larger than in our studies, which we believe is at the threshold of the DDR response. Therefore, we believe the damage being produced in our study is less than in the previous studies but above the threshold to activate the DDR. Our results help explain the previously reported inhibition of cytokinesis induced by laser microirradiation at chromosome ends.3 Here, we show that laser-induced DNA breaks (DSBs and SSBs) at a single chromosome end are capable of activating a DDR during anaphase onset via recruitment of γH2AX. Furthermore, proteins involved in the DDR such as PARP1, Nbs1, and cell cycle delay proteins, Chk1 and Chk2, are recruited to laser-induced DNA breaks at single chromosome ends as well as internal chromosome sites in anaphase cells (Table 1). However, only NIR focal DNA breaks at chromosome ends recruit phosphorylated p53 serine 15, whereas internal chromosomal breaks did not [Fig. 3(b), lane 3, ; Table 1]. It is unclear as to why p53 is not phosphorylated at internal chromosomal breaks; however, one possibility is a lack of full DDR in mitosis.30 In the case of the telomere-containing chromosome ends, this result may be consistent with the unique activity of the telomere as a protection from degradation processes, recombination, and chromosome fusion events. In addition, induced DNA breaks at chromosome ends and chromosome arms recruit endogenous NHEJ repair complex and PCNA during anaphase onset, suggesting a processing role in DNA repair [Figs. 4(a) and 4(b), lanes 1 and 2; Table 1], but DNA lesions at both chromosome ends and chromosome arms fail to recruit HR repair protein Rad51 [Table 1 and Fig. 5(a)]. Furthermore, our unpublished data suggest damaged chromosome ends continue into G1 phase with unresolved repair, as previously shown in interphase cells.10 In this study, we combine focal point femtosecond NIR laser microirradiation with immunofluorescence to understand whether additional DDR factors are recruited to DNA breaks at specific chromosome sites. This approach provides the opportunity to study DNA repair in single cells. Collectively, our data suggest that damaged anaphase chromosome ends and damaged chromosome arms activate DDR and may be processed by NHEJ based on the recruitment of protein complex. Our results also demonstrate that the inhibition of cytokinesis, as previously shown,3 is due to the activation of a DDR at laser-induced DNA breaks on single anaphase chromosome ends. These results are significant because of the cell’s ability to protect chromosome ends in order to prevent the activation of the DDR, which is consistent with the protective role of telomeres in maintaining genomic stability. Further studies should address the possible recruitment of additional DDR factors between damage chromosome regions and determine if there are differences in the kinetics of the DDR at both damage sites. 5.ConclusionA focal point 800-nm fs NIR microirradiation system can be used to study the effect of DNA break induction by the production of DSBs and SSBs at either a chromosome end or an internal chromosomal site during early anaphase of cells. Our results demonstrate that DNA breaks induced at either site are able to activate the DDR that results in the recruitment of cell cycle response factors Chk1, Chk2, and repair proteins . There appears to be uniqueness in the response of the chromosome ends in that they also recruit p53 phosphorylation on serine 15 (Ser15), whereas the damaged chromosome arms do not. Due to its high temporal and spatial resolution, laser microirradiation can be used to study the activation of the DDR at a single cell level, the effect of DNA breaks at different chromosome regions during mitosis and in subsequent stages of the cell cycle. AcknowledgmentsWe are grateful to Dr. Kyoko Yokomori (UC-Irvine) for kind donation of anti-PARP1, Dr. Jagesh V. Shah (Harvard Medical School) for helpful discussions, Cell Signaling Technology for anti-PCNA, and DAKO Company (Carpinteria, CA) for Cy3-congugated (TTAGGG)-PNA kit. This work was supported by the National Institutes of Health Laser Microbeam and Medical Program (RR01192), the Air Force Office of Scientific Research (FA9550-04-1-0101), Beckman Laser Institute Inc. Foundation (to M.W.B.), UCI-MBRS Program NIH Grant (GM055246, to B.A.S.), and Ford Foundation Fellowship from the National Academy of Sciences (to B.A.S.). ReferencesV. Gomez-Godinezet al.,
“Recruitment of DNA damage recognition and repair pathway proteins following near-IR femtosecond laser irradiation of cells,”
J. Biomed. Opt., 12
(2), 020505
(2007). http://dx.doi.org/10.1117/1.2717684 JBOPFO 1083-3668 Google Scholar
V. Gomez-Godinezet al.,
“Analysis of DNA double-strand break response and chromatin structure in mitosis using laser microirradiation,”
Nucleic Acids Res., 38
(22), e202
(2010). http://dx.doi.org/10.1093/nar/gkq836 NARHAD 0305-1048 Google Scholar
N. M. Bakeret al.,
“Chromosome tips damaged in anaphase inhibit cytokinesis,”
PLoS One, 5
(8), e12398
(2010). http://dx.doi.org/10.1371/journal.pone.0012398 1932-6203 Google Scholar
T. de Lange,
“Shelterin: the protein complex that shapes and safeguards human telomeres,”
Genes Dev., 19
(18), 2100
–2110
(2005). http://dx.doi.org/10.1101/gad.1346005 GEDEEP 0890-9369 Google Scholar
T. de Lange,
“Protection of mammalian telomeres,”
Oncogene, 21
(4), 532
–540
(2002). http://dx.doi.org/10.1038/sj.onc.1205080 ONCNES 0950-9232 Google Scholar
W. PalmT. de Lange,
“How shelterin protects mammalian telomeres,”
Annu. Rev. Genet., 42 301
–334
(2008). http://dx.doi.org/10.1146/annurev.genet.41.110306.130350 ARVGB7 0066-4197 Google Scholar
H. TakaiA. SmogorzewskaT. de Lange,
“DNA damage foci at dysfunctional telomeres,”
Curr. Biol., 13
(17), 1549
–1556
(2003). http://dx.doi.org/10.1016/S0960-9822(03)00542-6 CUBLE2 0960-9822 Google Scholar
M. T. Hayashiet al.,
“A telomere-dependent DNA damage checkpoint induced by prolonged mitotic arrest,”
Nat. Struct. Mol. Biol., 19
(4), 387
–394
(2012). http://dx.doi.org/10.1038/nsmb.2245 1545-9993 Google Scholar
W. B. Daltonet al.,
“Human cancer cells commonly acquire DNA damage during mitotic arrest,”
Cancer Res., 67
(24), 11487
–11492
(2007). http://dx.doi.org/10.1158/0008-5472.CAN-07-5162 CNREA8 0008-5472 Google Scholar
M. Fumagalliet al.,
“Telomeric DNA damage is irreparable and causes persistent DNA–damage–response activation,”
Nat. Cell Biol., 14
(4), 355
–365
(2012). http://dx.doi.org/10.1038/ncb2466 NCBIFN 1465-7392 Google Scholar
M. L. Duquetteet al.,
“CtIP is required to initiate replication-dependent interstrand crosslink repair,”
PLoS Genet., 8
(11), e1003050
(2012). http://dx.doi.org/10.1371/journal.pgen.1003050 PGLEB5 1553-7404 Google Scholar
X. Konget al.,
“Comparative analysis of different laser systems to study cellular responses to DNA damage in mammalian cells,”
Nucleic Acids Res., 37
(9), e68
(2009). http://dx.doi.org/10.1093/nar/gkp221 NARHAD 0305-1048 Google Scholar
J. S. Kimet al.,
“In situ analysis of DNA damage response and repair using laser microirradiation,”
Methods Cell Biol., 82 377
–407
(2007). http://dx.doi.org/10.1016/S0091-679X(06)82013-3 MCBLAG 0091-679X Google Scholar
D. Trautleinet al.,
“Specific local induction of DNA strand breaks by infrared multi-photon absorption,”
Nucleic Acids Res., 38
(3), e14
(2010). http://dx.doi.org/10.1093/nar/gkp932 NARHAD 0305-1048 Google Scholar
S. W. Botchwayet al.,
“Use of near infrared femtosecond lasers as sub-micron radiation microbeam for cell DNA damage and repair studies,”
Mutat. Res., 704
(1–3), 38
–44
(2010). http://dx.doi.org/10.1016/j.mrrev.2010.01.003 MUREAV 0027-5107 Google Scholar
A. Inagakiet al.,
“Dynamic localization of human RAD18 during the cell cycle and a functional connection with DNA double-strand break repair,”
DNA Repair (Amst), 8
(2), 190
–201
(2009). http://dx.doi.org/10.1016/j.dnarep.2008.10.008 DRNEAR 1568-7864 Google Scholar
P. O. Mariet al.,
“Dynamic assembly of end-joining complexes requires interaction between and XRCC4,”
Proc. Natl. Acad. Sci. U.S A., 103
(49), 18597
–18602
(2006). http://dx.doi.org/10.1073/pnas.0609061103 PNASA6 0027-8424 Google Scholar
J. S. Kimet al.,
“Specific recruitment of human cohesin to laser-induced DNA damage,”
J. Biol. Chem., 277
(47), 45149
–45153
(2002). http://dx.doi.org/10.1074/jbc.M209123200 JBCHA3 0021-9258 Google Scholar
E. Ferrando-Mayet al.,
“Highlighting the DNA damage response with ultrashort laser pulses in the near infrared and kinetic modeling,”
Front. Genet., 4 135(1–8)
(2013). http://dx.doi.org/10.3389/fgene.2013.00135 1664-8021 Google Scholar
J. R. Stoutet al.,
“Deciphering protein function during mitosis in PtK cells using RNAi,”
BMC Cell Biol., 7 26(1–15)
(2006). http://dx.doi.org/10.1186/1471-2121-7-26 BCBMAY 1471-2121 Google Scholar
C. L. RiederR. W. Cole,
“Entry into mitosis in vertebrate somatic cells is guarded by a chromosome damage checkpoint that reverses the cell cycle when triggered during early but not late prophase,”
J. Cell Biol., 142
(4), 1013
–1022
(1998). http://dx.doi.org/10.1083/jcb.142.4.1013 JCLBA3 0021-9525 Google Scholar
M. W. BernsD. E. RoundsR. S. Olson,
“Effects of laser micro-irradiation on chromosomes,”
Exp. Cell Res., 56
(2), 292
–298
(1969). http://dx.doi.org/10.1016/0014-4827(69)90016-0 ECREAL 0014-4827 Google Scholar
E. L. BotvinickM. W. Berns,
“Internet-based robotic laser scissors and tweezers microscopy,”
Microsc. Res. Tech., 68
(2), 65
–74
(2005). http://dx.doi.org/10.1002/(ISSN)1097-0029 MRTEEO 1059-910X Google Scholar
G. L. MoldovanB. PfanderS. Jentsch,
“PCNA, the maestro of the replication fork,”
Cell, 129
(4), 665
–679
(2007). http://dx.doi.org/10.1016/j.cell.2007.05.003 CELLB5 0092-8674 Google Scholar
D. A. SolomonM. C. CardosoE. S. Knudsen,
“Dynamic targeting of the replication machinery to sites of DNA damage,”
J. Cell Biol., 166
(4), 455
–463
(2004). http://dx.doi.org/10.1083/jcb.200312048 JCLBA3 0021-9525 Google Scholar
J. Esserset al.,
“Nuclear dynamics of PCNA in DNA replication and repair,”
Mol. Cell Biol., 25
(21), 9350
–9359
(2005). http://dx.doi.org/10.1128/MCB.25.21.9350-9359.2005 MCBDEU 1470-0573 Google Scholar
P. BaumannS. C. West,
“Role of the human RAD51 protein in homologous recombination and double-stranded-break repair,”
Trends Biochem. Sci., 23
(7), 247
–251
(1998). http://dx.doi.org/10.1016/S0968-0004(98)01232-8 TBSCDB 0167-7640 Google Scholar
S. E. Petersonet al.,
“Cdk1 uncouples CtIP-dependent resection and Rad51 filament formation during M-phase double-strand break repair,”
J. Cell Biol., 194
(5), 705
–720
(2011). http://dx.doi.org/10.1083/jcb.201103103 JCLBA3 0021-9525 Google Scholar
K. O. Greulich,
“Photons bring light into DNA repair: the comet assay and laser microbeams for studying photogenotoxicity of drugs and ageing,”
J. Biophotonics, 4
(3), 165
–171
(2011). http://dx.doi.org/10.1002/jbio.v4.3 JBOIBX 1864-063X Google Scholar
S. GiuntaR. BelotserkovskayaS. P. Jackson,
“DNA damage signaling in response to double-strand breaks during mitosis,”
J. Cell Biol., 190
(2), 197
–207
(2010). http://dx.doi.org/10.1083/jcb.200911156 JCLBA3 0021-9525 Google Scholar
|
||||||||||||||||||||||||||||||||||||||||||||||||



