

|
|
|
Vulnerable plaques (VPs) are metabolically active and histologically complex atheromas that are prone to spontaneous rupture often leading to sudden death.1 Coronary angiography cannot distinguish between a VP and a stable plaque. Mid-infrared (mid-IR) spectroscopy has been used previously to characterize the biochemical changes within thin sections of atheromas including VPs ex vivo. 2, 3, 4, 5 A variety of absorption peaks and bands within the mid-IR transmission spectra have been specifically linked to the vibrations of atoms within atherogenic lipoproteins, phospholipid particles, cholesterol esters, and fatty acids. Unfortunately, the high native water content within the arterial wall and its strong interfering absorption has severely limited the use of mid-IR spectroscopy. Alternative methods can circumvent these difficulties, including spectral analyses of the absorption patterns from “reflected” as opposed to “transmitted” infrared light, coupled to a bright but harmless light emitter such as a synchrotron.6 In this article, we describe how mid-IR light is reflected by pathologic components in the diseased aortic intima of ApoE(-/-) mice. The reflected light also contains the spectral signatures of the pathologic molecules within atheromas including VPs. Reflection-based mid-IR spectroscopy is a well-established technique, and due to its unique contrast mechanism, it has been widely used for the characterization and structure-property elucidation of multilayered materials.7 We have previously extended this technique to measure the reflection-absorption Fourier transform infrared spectra (FTIR) of microbial communities located on geological materials that contain multiple reflective interfaces of different optical properties.6 We postulated that this technique could also be applied to whole tissues in which a subsurface change in refractive index creates a reflective interface among the pathologic molecules of interest. This type of interface exists within the intima of atherosclerotic vessels where non-native lipid (average refractive indicies of 1.45 to 1.50, depending on composition and sizes of lipid particles8) and calcium deposits like hydroxylapatite or calcium phosphate (average refractive index of at least 1.639) are found in great abundance. Normal intimal tissues are water content, relatively uniform in optical properties, and have an average refractive index of approximately 1.35 to 1.38 (by extrapolation from the literature).10, 11 Although these refractive indices are for visible light, they may approximate the differences within the complex atherosclerotic matrix. Conceptually, mid-IR light that enters an atheroma should encounter multiple refractive boundaries, wherein some light might be reflected and reemerge. From first principles, if the surface roughness at the refractive boundary is larger than the wavelength of the incoming light, the light that is not absorbed would undergo multiple diffuse reflections before reemerging. If the surface roughness is smaller than the wavelength (nearly smooth), unabsorbed light can experience a quasi-specular reflection before reemerging. The lack of information about the wavelength-dependent refractive index, absorption coefficients, and the surface roughness of atherosclerotic interfaces makes it difficult to predict reflectivity. Given the morphologic complexity of most atheromas, the final signal would likely be the sum of these two modes of reflectivity. It is also likely that the features of the reflected infrared spectra would contain the absorption patterns generated by the major pathologic components of an atheroma, namely, atherogenic lipoproteins, extracellular phospholipid particles, smooth muscle cells, foam cells, and disintegrating foam cells as they undergo apoptotic cell death.12 We made infrared measurements on the explanted aorta of a 9-month-old female adult ApoE(-/-) mouse. ApoE(-/-) ( ) and wild-type mice were obtained from Jackson Laboratories (Bar Harbor, Maine). ApoE(-/-) mice were placed on a high fat/cholesterol (1 to 2%) diet for 3 months until sacrifice. We employed Beamline 1.4.3 at the advanced light source (ALS) synchrotron at LBNL (http://infrared.als.lbl.gov/). The synchrotron infrared beam has brightness of approximately to mrad-0.1%BW when focused to a spot. Specimens were placed luminal side up beneath an infrared-transparent window (International Crystal Labs, New Jersy) inside a custom-built environmental chamber kept at to preserve the “freshness” of each specimen. FTIR measurements were recorded in the standard reflection mode in the mid-IR 4,000 to region, and consisted of 128 co-added spectra at a spectral resolution of . We found that a highly focused beam (10 to ) was needed to avoid the interference of water from nondiseased regions and that a focal spot of or greater could not resolve plaque spectra separate from water signal. We also found that FTIR measurements could be obtained even through a layer of water. Figure 1 shows examples of the FTIR measurements from a cluster of atheromas including VPs (confirmed later by histologic examination) within the aortic arch. A significant number of mid-IR photons [Fig. 1a] that entered any given atheroma were reflected and emerged from the tissue sample [Fig. 1b]. In contrast, few were reflected from the nondiseased portions of aorta [Fig. 1c]. A striking feature was that the reflected light contained the spectral features [Fig. 1d] associated with atheromas.3, 4, 5 For instance, the 3100 to region revealed similar spectral features that arise from the carbon hydrogen bond stretching vibrations of fatty acids and cholesterol esters, including the moiety of their unsaturated hydrocarbon chains at or from their acyl groups at and . The strong absorption at also matched the spectral profile of the stretching vibrations of ester carbonyl groups of atherogenic phospholipid. The broader absorptions in the 1700 to region had comparable features to those generated by the stretching of the amide I and of the bending of the amide II modes present in proteins. In the 1500 to fingerprint region, absorption characteristics at and were similar to the known vibrations of the lipid acyl and to the symmetric bending of the lipid methyl groups, respectively. The absorptions at and matched the spectral patterns that arise from the asymmetric and symmetric stretching modes of in the phosphodiester groups of phospholipids, whereas absorptions centered at and matched the ester vibrations of phospholipid, cholesterol ester and fatty acid.3, 4, 5 Fig. 1Typical intensity profiles of (a) the incoming synchrotron infrared beam, (b) reflected signals from atheromas, and (c) nonatherosclerotic sites. (d) The corresponding reflection-absorption spectra of (b) shared spectral characteristics that were consistent with known excitation effects by infrared photons on atoms of molecules that compose atheromas. Each plot shows the averaged spectrum (black line) standard deviation (gray line); . 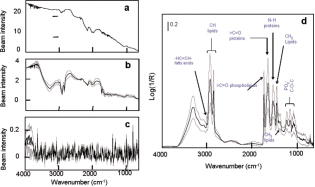 A dendrographic analysis of all FTIR measurements (employing -value distances measure, Ward’s algorithm, together with the distribution of the values, not shown) suggested that our spectra data could be grouped into four categories (I to IV) [Figs. 2a, 2b, 2c, 2d ; black lines], based on their similarity. To determine whether the observed spectral patterns of reflected light could be linked to particular pathologic features, we conducted additional FTIR measurements on several model systems, (Fig. 3 ) & [Figs. 2a, 2b, 2c, 2d; gray lines]. Fig. 2(a) Spectral variations in the type-I spectrum of atheromas (black line) can be explained (see text) by the two-component model spectrum of mixed AL+PP (gray line). (b) Spectral variations in the type-II spectrum of atheromas (black line) can be explained (see text) by the three-component model spectrum of mixed (gray line). (c) Spectral variations in the type-III spectrum of atheromas (black line) can be explained (see text) by the four-component model spectrum of AL+PP+FC+SMC (gray line). Inset: a close-up comparison of the type-III spectrum of atheromas (black line) with the composite spectrum of (gray line). The triplet of peaks in 1300 to and the doublet of peaks in 1100 to closely match the typical spectral pattern of type-I collagen (see text). (d) Spectral variations in the type-IV spectrum of atheromas (black line) can be explained (see text) by the spectrum of DFC (gray line). R, reflectance; AL, atherogenic lipoproteins; PP, phospholipid particles; AM, activated macrophages; FC, late-stage foam cells; DFC disintegrating foam cells, and SMC, smooth muscle cells. 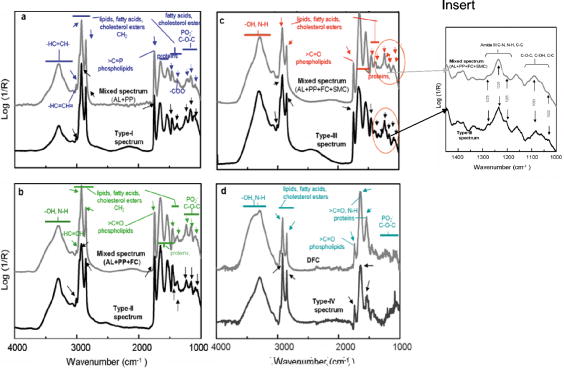 Fig. 3Photomicrographs and reflection-absorption FTIR spectra of AL, PP, AM, FC, SMC, and DFC are displayed. Each spectrum is an average of at least 20 different samples. Purified chicken egg yolk and lipopolysaccharides from Salmonella minnesota were purchased from Sigma-Aldrich (Missouri). J744A.1 murine macrophage cells were obtained from the American Tissue Culture Collection (ATCC). Thin films of egg yolk14 were prepared from the powder of the purified egg yolk, which was suspended in distilled sterile water and dried under a stream of nitrogen gas. We recorded the spectra of extracellular PL and SMC that were isolated from transverse microdissections of frozen ApoE(-/-) mouse tissues. To obtain spectra of macrophage and macrophage-derived foam cells, we activated mouse macrophages (J744A.1) in culture with bacterial molecules and obtained foam cells by feeding the activated macrophages cholesterol-containing liposomes. The spectra of each stage of foam cell evolution were measured at (early-stage cells), (mature), and (late). For DFCs, we fed activated macrophages excess cholesterol-containing liposomes for . Scale bars PP, ; AM, FC, and DFC, ; and SMC, . 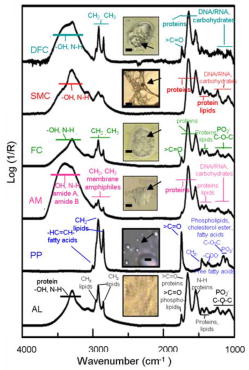 We found that the type-I spectrum [Fig. 2a, black lines], based on its spectral shape and the sharply defined high absorptions (at , , , , , , , , and ), indicates the dominant presence of lipoproteins and phospholipids [Fig. 3a; gray line]. For the type-II spectrum [Fig. 2b; black line], its overall spectral shape indicates the presence of foam cells [Fig. 2b; gray line] in addition to lipoprotein and phospholipid; a characteristic of VPs. The type-III spectrum exhibits a fine structure of a triplet of peaks at , , and , and a doublet of peaks at and [Fig. 2c; black line], are indicative of type-I collagen3, 5 and possibly smooth muscle cells [Fig.2c; gray line]. Smooth muscle cells in VPs that have been reported to orchestrate the assembly of type-I collagen.13 Finally, the type-IV spectrum [Fig. 2d; black line] exhibits spectral features in the region typical of the disintegrating foam cells [Fig. 2d; gray line], especially the spectral region between 1200 and , in which there was a conspicuous absence of the group absorption peaks that are characteristic of nucleic acids and various oligo- and polysaccharides, most of which were probably degraded during apoptotic cell death. We then repeated FTIR measurements on the diseased aortas from another three ApoE(-/-) mice (3-month-old adult females) and two wild-type controls and found similar results. Further studies are required to determine whether these new findings can be translated to the diseased vessels of other animal models or humans. ReferencesM. Gossl,
“Vulnerable plaque: detection and management,”
Med. Clin. North Am., 91
(4), 573
–601
(2007). 0025-7125 Google Scholar
J. M. Gentner,
E. Wentrup-Byrne,
P. J. Walker, and
M. D. Walsh,
“Comparison of fresh and post-mortem human arterial tissue: an analysis using FT-IR microspectroscopy and chemometrics,”
Cell. Mol. Biol. (Paris), 44
(1), 251
–259
(1998). 0145-5680 Google Scholar
K. Z. Liu,
I. M. C. Dixon, and
H. H. Mantsch,
“Distribution of collagen deposition in cardiomyopathic hamster hearts determined by infrared microscopy,”
Cardiovasc. Pathol., 8
(1), 41
–47
(1999). 1054-8807 Google Scholar
A. Becker,
M. Epple,
K. M. Muller, and
I. Schmitz,
“A comparative study of clinically well-characterized human atherosclerotic plaques with histological, chemical, and ultrastructural methods,”
J. Inorg. Biochem., 98
(12), 2032
–2038
(2004). 0162-0134 Google Scholar
D. L. Wetzel,
G. R. Post, and
R. A. Lodder,
“Synchrotron infrared micro spectroscopic analysis of collagens I, III, and elastin on the shoulders of human thin-cap fibroatheromas,”
Vib. Spectrosc., 38
(1–2), 53
–59
(2005). 0924-2031 Google Scholar
H.-Y. N. Holman and
M. C. Martin,
“Synchrotron radiation infrared spectromicroscopy: a non-invasive molecular probe for biogeochemical processes,”
Advances in Agronomy, 79
–127 Elsevier, New York (2006). Google Scholar
M. Claybourn,
“Mid-infrared external reflection spectroscopy,”
Handbook of Vibrational Spectroscopy, 969
–981 Wiley Interscience, London (2002). Google Scholar
V. P. Maltsev,
A. V. Chernyshev,
K. A. Semyanov, and
E. Soini,
“Absolute real-time determination of size and refractive index of individual microspheres,”
Meas. Sci. Technol., 8
(9), 1023
–1027
(1997). https://doi.org/10.1088/0957-0233/8/9/011 0957-0233 Google Scholar
B. J. Tarasevich,
C. C. Chusuei, and
D. L. Allara,
“Nucleation and growth of calcium phosphate from physiological solutions onto self-assembled templates by a solution-formed nucleus mechanism,”
J. Phys. Chem. B, 107
(38), 10367
–10377
(2003). 1089-5647 Google Scholar
L. J. J. Dirckx,
L. C. Kuypers, and
W. F. Decraemer,
“Refractive index of tissue measured with confocal microscopy,”
J. Biomed. Opt., 10
(4), 044014
(2005). https://doi.org/10.1117/1.1993487 1083-3668 Google Scholar
H. D. Downing and
D. Williams,
“Optical-constants of water in infrared,”
J. Geophys. Res., 80
(12), 1656
–1661
(1975). 0148-0227 Google Scholar
P. Libby and
P. Theroux,
“Pathophysiology of coronary artery disease,”
Circulation, 111
(25), 3481
–3488
(2005). 0009-7322 Google Scholar
M. M. Kockx,
“Cell composition, replication, and apoptosis inatherosclerotic plaques after 6 months of cholesterol withdrawal,”
Circ. Res., 83
(4), 378
–387
(1998). 0009-7330 Google Scholar
D. Kritchevsky,
“Dietary-protein, cholesterol and atherosclerosis—a review of the early history,”
J. Nutr., 125
(3), S589
–S593
(1995). 0022-3166 Google Scholar
|



