

|
|
1.IntroductionHydrogen peroxide (), a common reactive oxygen species (ROS) found in biological systems, is now recognized as an intracellular second messenger for cellular signaling that exerts diverse physiological and pathological effects.1–7 The aberrant production or accumulation of within cellular mitochondria over time due to oxidative stress or genetic mutation is connected to serious pathological conditions including cancer,8 diabetes,9 and neurodegenerative diseases such as Alzheimer’s, Parkinson’s, and Huntington’s diseases, as well as stroke.10–12 In addition, is involved in therapeutic processes such as wound healing, stem cell proliferation, and an adaptive response in astrocytes leading to neuronal protection.1,2,7,13 A substantial challenge in elucidating the diverse roles of in complex biological environments is the lack of methods to determine the spatial and temporal dynamics of this reactive oxygen metabolite in living systems. For the detection of ROS production in vitro, several fluorescent probes have been developed based on small molecules, fluorescent proteins, and nanoparticles.3,14–19 Among these technologies, small molecule probes offer an attractive approach to ROS detection due to their general compatibility with an array of biological systems without external activating enzymes and genetic manipulation. However, traditional small molecule probes such as dichlorofluorescein (DCF) derivatives detect multiple types of reactive small molecules, including other ROS such as superoxide radical (), hydroperoxy radical (), singlet oxygen (), peroxy radical (), and reactive nitrogen species;16 they are not specific for . In addition, DCF derivatives cannot be targeted to specific intracellular compartments.14 To overcome the disadvantages of existing methods for detecting ROS, new chemoselective fluorescent indicators featuring a boronate-based molecular detection mechanism have been developed,3 which provide improved selectivity for over related ROS, particularly superoxide, nitric oxide, and hydroxyl radical. These probes include peroxyfluor-2 (PF2), peroxy yellow 1 (PY1), peroxy orange 1 (PO1), peroxyfluor-6 acetoxymethyl ester (PF6-AM), mitochondria peroxy yellow 1 (MitoPY1), etc.2,3,18,20–22 However, conventional confocal microscopy has limitations for use in real time in vivo imaging, including photodamage, photobleaching, and limited imaging depth. Furthermore, prolonged visible light exposure can result in artifactual ROS generation and signal amplification.23,24 Therefore, two-photon imaging of offers an attractive alternative to overcome many of these limitations.25–27 We report two-photon fluorescence (TPF) imaging for the detection of intracellular cytoplasmic and mitochondrial production in live cells using a variety of next-generation boronate-based probes such as PF6-AM and MitoPY1. Two-photon absorption (TPA) cross sections of these probes were measured with a Ti:sapphire laser by comparison with the TPA cross section of fluorescein.26,28 To our knowledge, this is the first two-photon characterization of these particular chemoselective probes and provides a clear example for the detection of intracellular production with TPF imaging. 2.Materials and Methods2.1.Chemoselective Fluorescent ProbesWe characterized a series of previously reported chemoselective probes with three useful colors: green (PF2 and PF6-AM), yellow (PY1 and MitoPY1), and orange (PO1). Figure 1 illustrates reaction of with these probes, which are based on fluorescein/rhodamine derivatives. The aryl boronate to phenol chemical switch is utilized for selective detection of over other ROS. Upon reaction with , a highly fluorescent product is released and can be assayed by fluorescence imaging. To improve the cell membrane permeability and trap the probe in the cytosol, PF6-AM was modified from PF6 with acetoxymethyl ester groups.2,26 Upon penetration of the cell membrane, PF6-AM is deprotected by intracellular esterases, releasing the dianionic PF6 that is then trapped in the cytosol [Fig. 1(e)]. MitoPY1 [Fig. 1(d)] was derived from PY1 to include a combination of a boronate-based switch and a mitochondrial-targeting phosphonium moiety for the detection of localized to cellular mitochondria.18 2.2.TPA MeasurementA basic parameter to characterize a fluorophore for TPF imaging is the TPA cross section. TPA spectra were measured using a Ti:sapphire laser (Spectra Physics Mai Tai HP) operating at 720 to 1040 nm, 100 fs pulse width, and 80 MHz repetition rate. The scheme of experimental setup was described in Xu and Webb.28 Before the measurement, we verified that the TPF intensity increases with the square of the excitation power. TPF intensity is related to spatial dependence and temporal dependence such as laser parameters and system collection condition.28 A direct measurement of TPA cross section is difficult. Therefore, absolute TPA cross section values of fluorescent probes () were calculated by comparison with the TPF intensity of fluorescein () in Eq. (1):28 where and are the fluorescence intensity measured with a photon counting photomultiplier tube (PMT) at the same laser power excitation, and are the two-photon-excited fluorescence quantum yields of the probes assumed to be same to that of single-photon excited, and are the concentrations of the probes, and are the quantum efficiency of the PMT (Hamamatsu R7600U-200) obtained from the manufacturer’s data. The system collection efficiency, estimated to be the same for the probes, are based on its fluorescence emission spectrum, the measured numerical aperture (NA) and transmission of the objective lens, and the transmission of the filters.28 The fluorescent probes were measured with the same measurement conditions as fluorescein, therefore these measurements can be used to calculate TPA cross section without explicitly characterizing pulse shape by comparing measured values to those found in Ref. 28.PF2, PY1, PO1, MitoPY1, and DCF were diluted to form a 20 μM solution in phosphate buffered saline (PBS) buffer. We then added high concentrations of (100 μM) to ensure complete deprotection. PF6-AM (20 μM) in PBS buffer was incubated with both (100 μM) and esterase () from rabbit liver (Sigma–Aldrich #040566) to deprotect the boronate and the AM-esters. 2.3.Confocal and Two-photon Microscopyimaging was performed with a commercial laser scanning inverted microscope system (Zeiss 710NLO) including configuration both for confocal and TPF microscopy. This system was equipped with a 488-nm argon laser and a Ti:sapphire laser (Coherent Chameleon Vision II) with 690- to 1080-nm wavelength, 140-fs pulse width, and 80-MHz repetition rate. We operated the Ti:sapphire laser at 770 nm, a wavelength for TPF imaging of PF6-AM, MitoPY1, and Hoechst 33342. A objective (Zeiss Plan-Apochromat) was employed to focus the excitation laser beam onto cells and was also used for fluorescence collection into the PMTs. A prism-based 34-channel QUASAR detection unit was used for tunable spectral band-width collection without traditional band-pass filters. A 5% circulation and 37°C thermal chamber was used for live cell imaging. 2.4.Cell CultureAll animal procedures were performed according to protocols approved by the Institutional Animal Care and Use Committee of the Weill Medical College of Cornell University. The HT22 cell, an immortal neuroblast line originated from hippocampal neurons, and primary rat astrocytes were cultured in 35-mm Petri dishes with a cover-glass at the bottom. Primary astrocyte cultures were prepared from the cerebral cortices of Sprague-Dawley rat pups (P1-3) as described in Haskew-Layton et al.1 In brief, astrocyte cultures were grown for about 2 weeks until reaching confluency in minimal essential medium (Invitrogen) supplemented with 10% horse serum and penicillin plus streptomycin. Once confluent, the astrocytes were treated with 8 μM cytosine-d-arabinofuranoside (Ara-C), a mitotic inhibitor for , to kill off contaminating cells. The astrocytes were used for experiments at 2 to 3 weeks in culture. 2.5.Intracellular Productionis generally produced by the dismutation of mitochondrial superoxide or as a product of enzymatic activity. To mimic mitochondrial production, HT22 cells were treated with the complex I respiratory chain inhibitor rotenone.29 To generate cytoplasmic , we used an enzymatic method for intracellular production in astrocytes.1 Primary astrocytes were transduced with adenoviruses containing the cDNA for cytoplasmic d-amino acid oxidase (DAAO) for 4 days. DAAO oxidatively deaminates d-amino acids using flavin adenin denucleotide (FAD) as an electron acceptor. At the same time, DAAO uses molecular to oxidize FAD, during which time is produced as a byproduct. is therefore produced in a dose-dependent manner relative to the concentration of d-alanine added. Following DAAO-transduction, astrocytes were incubated with 5 μM PF6-AM and Hoechst 33342. d-Alanine (2 mM) stimulated production in DAAO astrocytes was then detected; the cells were supplemented with the DAAO cofactor FAD (2.5 μM). 3.Results and Discussions3.1.TPA SpectrumThe two-photon activation times, single-photon absorption and emission peaks, and quantum yields of the probes were measured and are shown in Table 1. Activation time refers to the average time when the TPF intensity increases to of the saturation intensity. This activation time is related to the deprotection efficiency. Compared with the commonly used nonspecific probe DCF, the chemoselective probes demonstrated much faster responses to . Table 1Optical parameters of H2O probes.
Figure 2 shows single-photon absorption and fluorescence emission spectra, and absolute values of TPA cross sections for the probes PF2, PY1, PO1, MitoPY1, and PF6-AM. As a comparison, Fig. 2(f) shows the TPA cross section of DCF. The peak cross section values of chemoselective probes are comparable to that of fluorescein,28 which is sufficiently large for two-photon imaging in vitro or in vivo. To ensure that has deprotected the boronate, the TPF spectra were measured at least 1 h after addition. Then, the TPA cross section was calculated based on the above Eq. (1) and the parameter in Table 1. Fig. 2Single-photon absorption spectrum (black line), fluorescence emission spectrum (green-dot line), and absolute value of two-photon absorption spectrum (red-symbol line) for probes in phosphate buffered saline solution. . (a) PF2, (b) PY1, (c) PO1, (d) MitoPY1, (e) PF6-AM, and (f) DCF.  We measured several color probes here so that multicolor imaging can be performed with minimum fluorescence bleed-through with other co-staining dyes. Figure 2 helps us to select co-staining dyes for single excitation wavelength two-photon multicolor imaging. 3.2.TPF Imaging of IntracellularFigure 3 shows intracellular TPF imaging of in HT22 cells stained with PF6-AM. The cells were incubated in PF6-AM solution for 20 min and the nuclei were stained with Hoechst 33342. Figures 3(a) and 3(b) show the concentration increasing in cells after the addition of exogenous . This result demonstrates that PF6-AM is capable of detecting cytoplasmic . Figures 3(c) and 3(d) demonstrate endogenous production induced by rotenone, which resulted in bright punctate staining patterns that are likely the sources of production. Fig. 3Two-photon fluorescence (TPF) imaging of extra additional and endogenous in HT22 cells. TPF imaging in (a) 1 min and (b) 38 min after extra 50 μM was added. TPF imaging in (c) 3 min and (d) 38 min after rotenone was added. The nuclei (blue), stained with Hoechst 33342, were co-excited with 770-nm wavelength laser pulses. Scale bar: 50 μm. 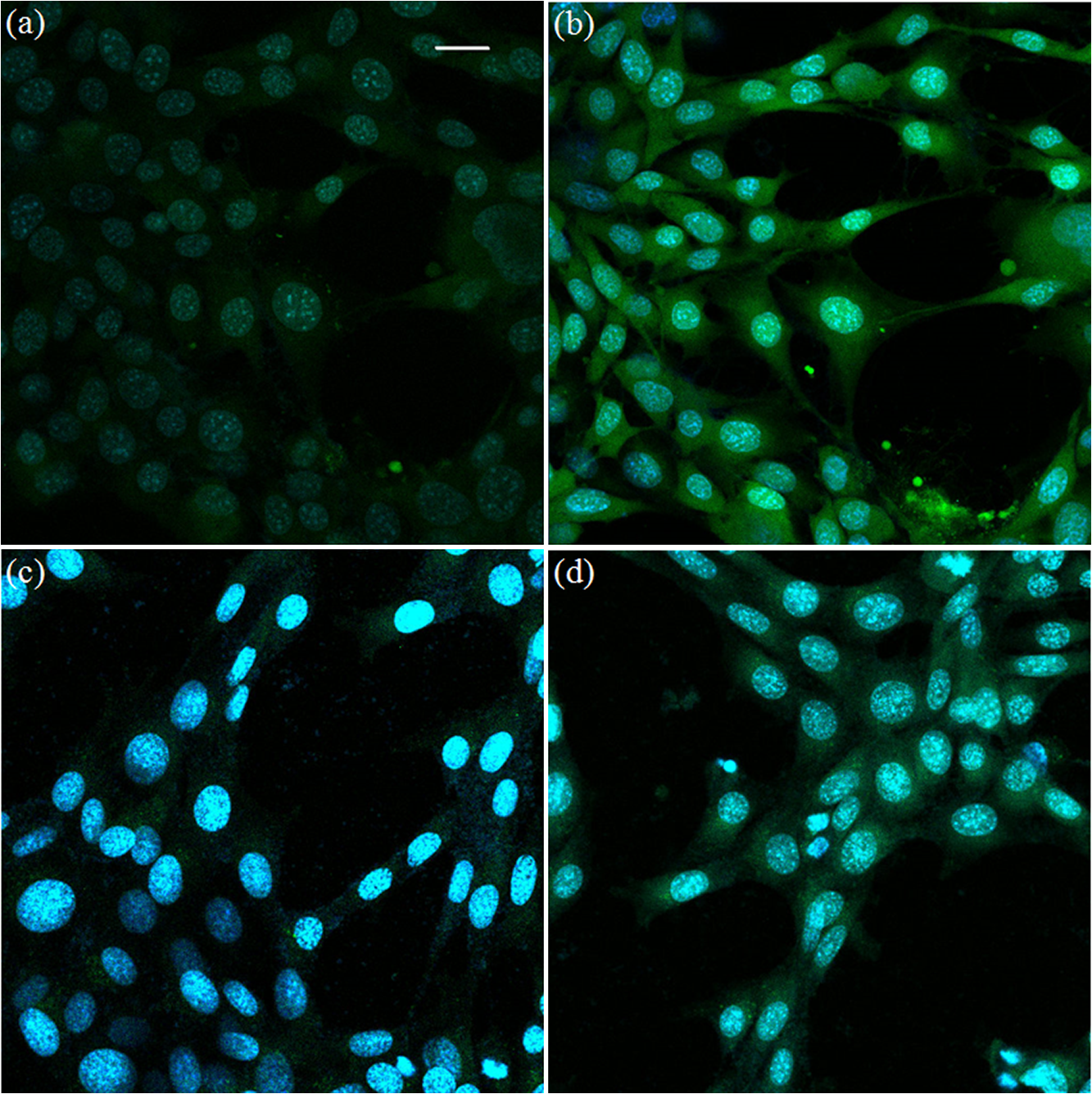 To improve the signal-to-background ratio and target mitochondrial production, we used MitoPY1 for mitochondrial labeling and detection of localized production. Figure 4 shows an overlay of TPF imaging of MitoPY1 labeled mitochondrial production and MitoTracker Red labeled mitochondria. Rotenone was added to the HT22 cells after they were incubated with 5 μM MitoPY1 for 25 min. Then, MitoTracker Red was added after another 60 min to ensure that MitoPY1 was targeted to the mitochondria in HT22 cells. Fig. 4TPF imaging of localized mitochondrial in HT22 cells with MitoPY1. (a) TPF imaging of HT22 cells after MitoPY1 was incubated for 10 min. (b) TPF imaging of the cells after rotenone was added for 10 min. (c) TPF imaging shows the overlay of co-localization mitochondrial production and cellular mitochondria after rotenone was added for 80 min. The green channel shows MitoPY1 signal (d) and the red channel shows MitoTracker Red signal (e). Both the MitoPY1 and MitoTracker Red dyes were co-excited with 770-nm wavelength laser pulses. Scale bar: 10 μm. 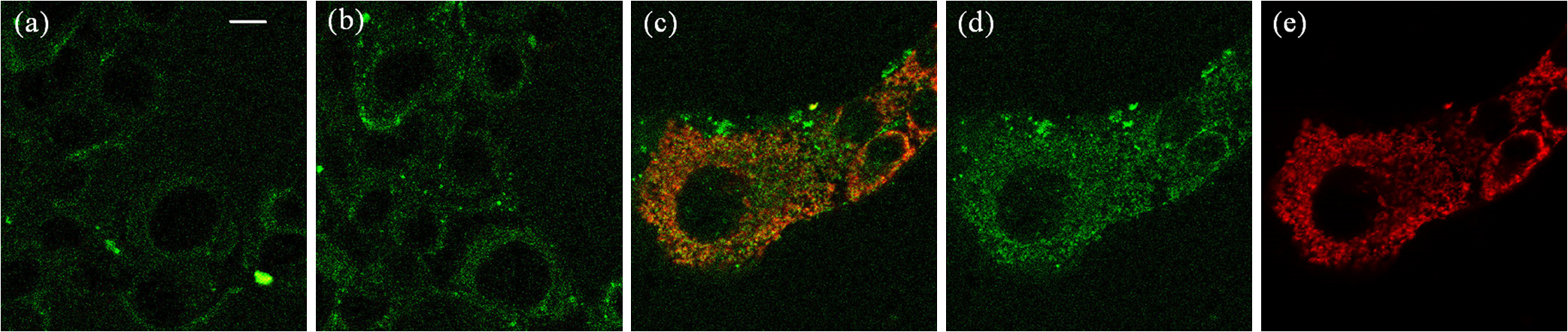 Figure 5 shows TPF imaging of cytoplasmic production induced by DAAO in astrocytes. PF6-AM and Hoechst 33342 were co-excited with 770-nm laser pulses. The TPA cross section of FAD at 770 nm is only that of PF6AM.30 Therefore, the fluorescence of FAD is only a weak background signal. The fluorescence intensity in Fig. 5 shows cytoplasmic increasing from the time (a) 1 min, (b) 6 min, and (c) 25 min after the addition of d-alanine and FAD. Fig. 5TPF images of cytoplasmic generation in astrocytes after (a) 1 min, (b) 6 min, and (c) 25 min after injection of d-alanine and FAD into the cells genetically modified with DAAO. The cells were loaded with 5 μM PF6-AM (green) for 30 min before use. The nuclei (blue), stained with Hoechst 33342, were co-excited with 770-nm wavelength laser pulses. Scale bar: 50 μm.  3.3.DiscussionThe involvement of in cellular signaling related to cancer and neurodegenerative diseases has motivated the development of imaging technologies for measuring intracellular concentration and dynamics in cellular compartments. Previous studies have relied on a horseradish peroxidase/Amplex Red substrate system to measure extracellular production using a spectrophotometer (Spectramax Plus 384; Molecular Devices).1 However, DAAO derived production provides a controllable scale ideal for intracellular measurements. TPF signal intensity is linearly related to the fluorophore concentration. Therefore, TPF imaging provides the possibility for quantification of localized intracellular production. The real-time visualization of production is another challenging issue that required a fast response of the fluorescent probes. Here, the chemoselective probes showed much faster activation time compared with commercial probes such as DCF. It provides a more accurate technique for flow cytometric analysis of isolated cells or mitochondria and allows for the detection of changes such as cell activation, oxidative status, and cell death. Some probes have been successfully used to study ROS in a variety of biological systems with confocal microscopy.31–34 Therefore, the combination of new fluorescent probes for and the deep tissue imaging capability of two-photon microscopy may allow direct visualization of in vivo dynamics in cells in their natural environment as well as their response to systematic manipulations. 4.ConclusionsThis study introduces chemoselective probes for TPF imaging intracellular production. TPA spectra were measured and TPF imaging was successfully demonstrated for cytoplasmic and mitochondrial production in brain cells using PF6-AM and MitoPY1 probes. The probes showed high sensitivity and fast response to detection. With the advantages of chemoselective probes, TFP imaging provides a novel opportunity for real-time monitoring of detection and oxidative stress evaluation in live cells and in vivo. AcknowledgmentsThis work is supported by the National Institutes of Health (NIH) Grant Nos. 2P01AG014930, 3R01 CA133148, R21AG042700, R21DK088066, R21EB012215, Dr. Miriam and Sheldon G. Adelson Medical Research Foundation. B.C.D. is a Fellow of the Jane Coffin Childs Memorial Fund for Medical Research. We acknowledge Christopher J. Chang, Demirhan Kobat, David Rivera, Anthony Fouad, Li Barie, Jian Zhong, and Jimmy B. Payappilly for technical support and helpful comments. ReferencesR. E. Haskew-Laytonet al.,
“Controlled enzymatic production of astrocytic hydrogen peroxide protects neurons from oxidative stress via an Nrf2-independent pathway,”
Proc. Natl. Acad. Sci. U.S.A., 107
(40), 17385
–17390
(2010). http://dx.doi.org/10.1073/pnas.1003996107 PNASA6 0027-8424 Google Scholar
B. C. Dickinsonet al.,
“Nox2 redox signaling maintains essential cell populations in the brain,”
Nat. Chem. Biol., 7
(2), 106
–112
(2011). http://dx.doi.org/10.1038/nchembio.497 1552-4450 Google Scholar
E. W. Milleret al.,
“Molecular imaging of hydrogen peroxide produced for cell signaling,”
Nat. Chem. Biol., 3
(5), 263
–267
(2007). http://dx.doi.org/10.1038/nchembio871 1552-4450 Google Scholar
S. G. Rhee,
“, a necessary evil for cell signaling,”
Science, 312
(5782), 1882
–1883
(2006). http://dx.doi.org/10.1126/science.1130481 SCIEAS 0036-8075 Google Scholar
B. C. DickinsonC. J. Chang,
“Chemistry and biology of reactive oxygen species in signaling or stress responses,”
Nat. Chem. Biol., 7
(8), 504
–511
(2011). http://dx.doi.org/10.1038/nchembio.607 1552-4450 Google Scholar
M. P. Murphyet al.,
“Unraveling the biological roles of reactive oxygen species,”
Cell Metab., 13
(4), 361
–366
(2011). http://dx.doi.org/10.1016/j.cmet.2011.03.010 CMEEB5 1550-4131 Google Scholar
P. Niethammeret al.,
“A tissue-scale gradient of hydrogen peroxide mediates rapid wound detection in zebrafish,”
Nature, 459
(7249), 996
–U123
(2009). http://dx.doi.org/10.1038/nature08119 NATUAS 0028-0836 Google Scholar
T. FinkelM. SerranoM. A. Blasco,
“The common biology of cancer and ageing,”
Nature, 448
(7155), 767
–774
(2007). http://dx.doi.org/10.1038/nature05985 NATUAS 0028-0836 Google Scholar
J. B. Piet al.,
“Reactive oxygen species as a signal in glucose-stimulated insulin secretion,”
Diabetes, 56
(7), 1783
–1791
(2007). http://dx.doi.org/10.2337/db06-1601 DIAEAZ 0012-1797 Google Scholar
K. J. BarnhamC. L. MastersA. I. Bush,
“Neurodegenerative diseases and oxidative stress,”
Nat. Rev. Drug Discov., 3
(3), 205
–214
(2004). http://dx.doi.org/10.1038/nrd1330 NRDDAG 1474-1776 Google Scholar
M. T. LinM. F. Beal,
“Alzheimer’s APP mangles mitochondria,”
Nat. Med., 12
(11), 1241
–1243
(2006). http://dx.doi.org/10.1038/nm1106-1241 1078-8956 Google Scholar
A. V. RaoB. Balachandran,
“Role of oxidative stress and antioxidants in neurodegenerative diseases,”
Nutr. Neurosci., 5
(5), 291
–309
(2002). http://dx.doi.org/10.1080/1028415021000033767 NNINFE 1476-8305 Google Scholar
C.-H. A. Chen,
“Role of reactive oxygen species in low level light therapy,”
Proc. SPIE, 7165 716502
(2009). http://dx.doi.org/10.1117/12.814890 PSISDG 0277-786X Google Scholar
V. V. Belousovet al.,
“Genetically encoded fluorescent indicator for intracellular hydrogen peroxide,”
Nat. Methods, 3
(4), 281
–286
(2006). http://dx.doi.org/10.1038/nmeth866 1548-7091 Google Scholar
B. Simen Zhaoet al.,
“A highly selective fluorescent probe for visualization of organic hydroperoxides in living cells,”
J. Am. Chem. Soc., 132
(48), 17065
–17067
(2010). http://dx.doi.org/10.1021/ja1071114 JACSAT 0002-7863 Google Scholar
J. P. Crow,
“Dichlorodihydrofluorescein and dihydrorhodamine 123 are sensitive indicators of peroxynitrite in vitro: implications for intracellular measurement of reactive nitrogen and oxygen species,”
Nitric Oxide, 1
(2), 145
–157
(1997). http://dx.doi.org/10.1006/niox.1996.0113 NIOXF5 1089-8603 Google Scholar
D. Lee et al.,
“In vivo imaging of hydrogen peroxide with chemiluminescent nanoparticles,”
Nat. Mater., 6
(10), 765
–769
(2007). http://dx.doi.org/10.1038/nmat1983 NMAACR 1476-1122 Google Scholar
B. C. DickinsonC. J. Chang,
“A targetable fluorescent probe for imaging hydrogen peroxide in the mitochondria of living cells,”
J. Am. Chem. Soc., 130
(30), 9638
–9639
(2008). http://dx.doi.org/10.1021/ja802355u JACSAT 0002-7863 Google Scholar
L. N. WuX. J. ZhangH. X. Ju,
“Highly sensitive flow injection detection of hydrogen peroxide with high throughput using a carbon nanofiber-modified electrode,”
Analyst, 132
(5), 406
–408
(2007). http://dx.doi.org/10.1039/b701933e ANLYAG 0365-4885 Google Scholar
A. R. LippertG. C. Van de BittnerC. J. Chang,
“Boronate oxidation as a bioorthogonal reaction approach for studying the chemistry of hydrogen peroxide in living systems,”
Acc. Chem. Res., 44
(9), 793
–804
(2011). http://dx.doi.org/10.1021/ar200126t ACHRE4 0001-4842 Google Scholar
J. ChanS. C. DodaniC. J. Chang,
“Reaction-based small-molecule fluorescent probes for chemoselective bioimaging,”
Nat. Chem., 4
(12), 973
–984
(2012). http://dx.doi.org/10.1038/nchem.1500 1755-4330 Google Scholar
B. C. DickinsonC. HuynhC. J. Chang,
“A palette of fluorescent probes with varying emission colors for imaging hydrogen peroxide signaling in living cells,”
J. Am. Chem. Soc., 132
(16), 5906
–5915
(2010). http://dx.doi.org/10.1021/ja1014103 JACSAT 0002-7863 Google Scholar
P. E. Hockbergeret al.,
“Activation of flavin-containing oxidases underlies light-induced production of in mammalian cells,”
Proc. Natl. Acad. Sci. U. S. A., 96
(11), 6255
–6260
(1999). http://dx.doi.org/10.1073/pnas.96.11.6255 PNASA6 0027-8424 Google Scholar
J. M. Squirrellet al.,
“Long-term two-photon fluorescence imaging of mammalian embryos without compromising viability,”
Nat. Biotechnol., 17
(8), 763
–767
(1999). http://dx.doi.org/10.1038/11698 NABIF9 1087-0156 Google Scholar
C. Chunget al.,
“A two-photon fluorescent probe for ratiometric imaging of hydrogen peroxide in live tissue,”
Chem. Commun., 47
(34), 9618
–9620
(2011). http://dx.doi.org/10.1039/c1cc13583j CHCOFS 1364-548X Google Scholar
H. Guoet al.,
“Two-photon imaging of intracellular hydrogen peroxide with a chemoselective fluorescence probe,”
Optics in the Life Sciences, Optical Society of America, Monterey, California
(2011). Google Scholar
Y. Chenet al.,
“Recent advances in two-photon imaging: technology developments and biomedical applications,”
Chin. Opt. Lett., 11
(1), 011703
(2013). http://dx.doi.org/10.3788/COL COLHBT 1671-7694 Google Scholar
C. XuW. W. Webb,
“Measurement of two-photon excitation cross sections of molecular fluorophores with data from 690 to 1050 nm,”
J. Opt. Soc. Am. B, 13
(3), 481
–491
(1996). http://dx.doi.org/10.1364/JOSAB.13.000481 JOBPDE 0740-3224 Google Scholar
N. Y. Liet al.,
“Mitochondrial complex I inhibitor rotenone induces apoptosis through enhancing mitochondrial reactive oxygen species production,”
J. Biol. Chem., 278
(10), 8516
–8525
(2003). http://dx.doi.org/10.1074/jbc.M210432200 JBCHA3 0021-9258 Google Scholar
S. H. HuangA. A. HeikalW. W. Webb,
“Two-photon fluorescence spectroscopy and microscopy of NAD(P)H and flavoprotein,”
Biophys. J., 82
(5), 2811
–2825
(2002). http://dx.doi.org/10.1016/S0006-3495(02)75621-X BIOJAU 0006-3495 Google Scholar
J. Luet al.,
“S100B and APP promote a gliocentric shift and impaired neurogenesis in down syndrome neural progenitors,”
PLoS One, 6
(7), e22126
(2011). http://dx.doi.org/10.1371/journal.pone.0022126 1932-6203 Google Scholar
Y. Ohsakiet al.,
“Increase of sodium delivery stimulates the mitochondrial respiratory chain production in rat renal medullary thick ascending limb,”
Am. J. Physiol-Renal, 302
(1), F95
–F102
(2012). http://dx.doi.org/10.1152/ajprenal.00469.2011 NEPSEY 0886-1714 Google Scholar
J. F. Woolleyet al.,
“ production downstream of FLT3 is mediated by p22phox in the endoplasmic reticulum and is required for STAT5 signalling,”
PLoS One, 7
(7), e34050
(2012). http://dx.doi.org/10.1371/journal.pone.0034050 1932-6203 Google Scholar
J. Sakaiet al.,
“Reactive oxygen species-induced actin glutathionylation controls actin dynamics in neutrophils,”
Immunity, 37
(6), 1037
–1049
(2012). http://dx.doi.org/10.1016/j.immuni.2012.08.017 IUNIEH 1074-7613 Google Scholar
|




